

Synergistic activation by Glass and Pointed promotes neuronal identity in the Drosophila eye disc
- PMID: 39154080
- PMCID: PMC11330500
- DOI: 10.1038/s41467-024-51429-z
Synergistic activation by Glass and Pointed promotes neuronal identity in the Drosophila eye disc
Abstract
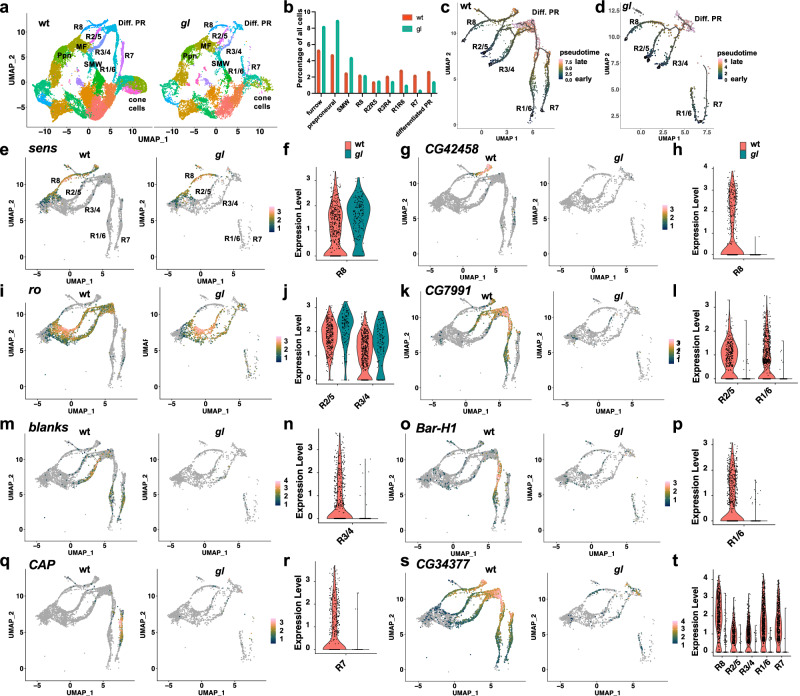
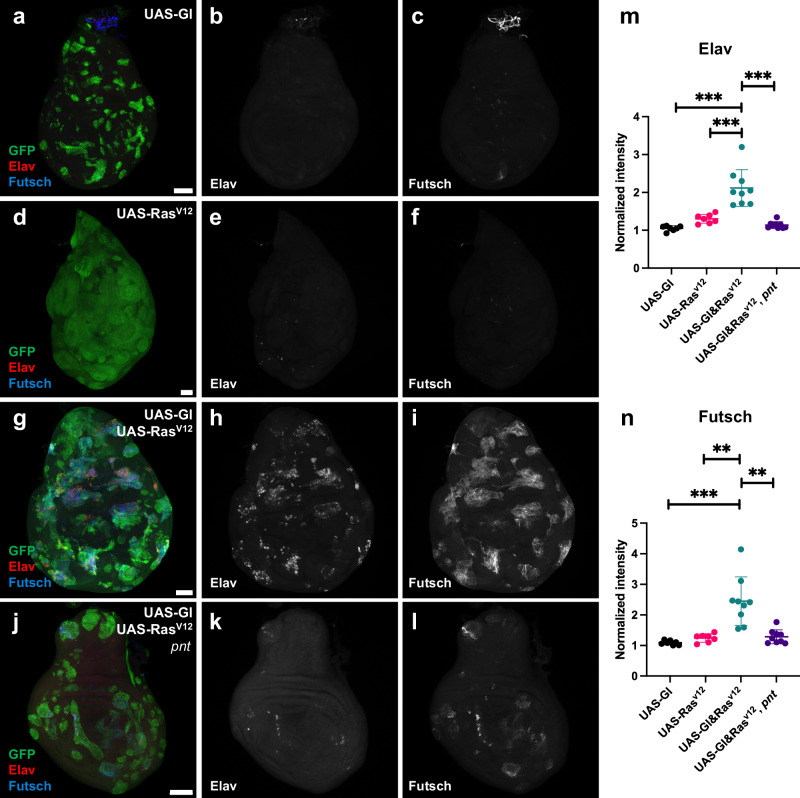
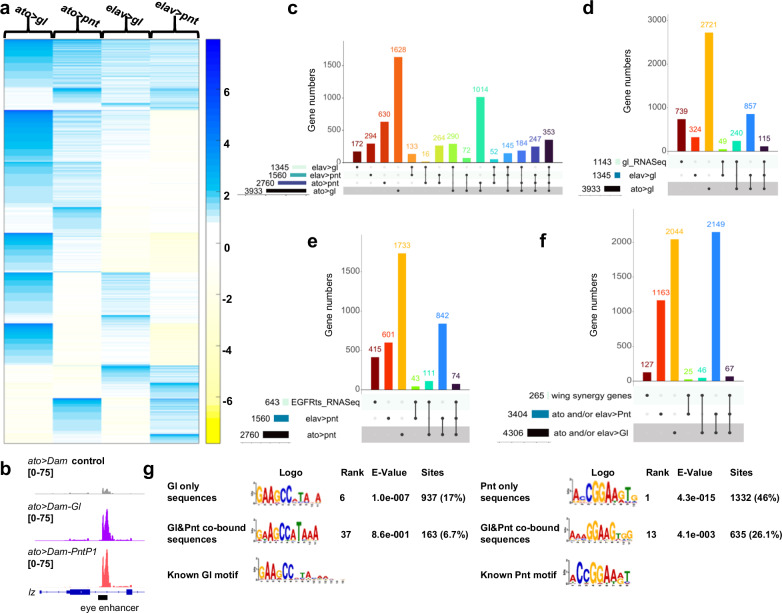
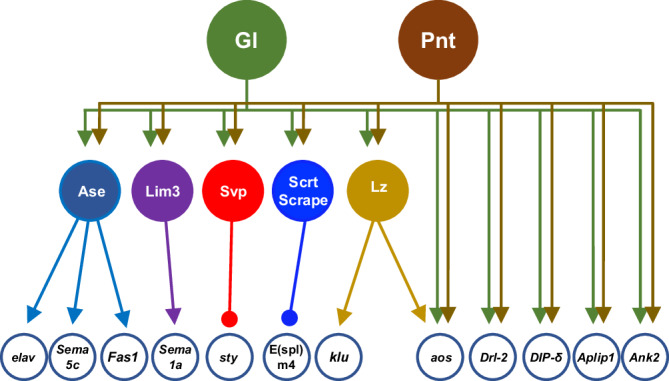
The integration of extrinsic signaling with cell-intrinsic transcription factors can direct progenitor cells to differentiate into distinct cell fates. In the developing Drosophila eye, differentiation of photoreceptors R1-R7 requires EGFR signaling mediated by the transcription factor Pointed, and our single-cell RNA-Seq analysis shows that the same photoreceptors require the eye-specific transcription factor Glass. We find that ectopic expression of Glass and activation of EGFR signaling synergistically induce neuronal gene expression in the wing disc in a Pointed-dependent manner. Targeted DamID reveals that Glass and Pointed share many binding sites in the genome of developing photoreceptors. Comparison with transcriptomic data shows that Pointed and Glass induce photoreceptor differentiation through intermediate transcription factors, including the redundant homologs Scratch and Scrape, as well as directly activating neuronal effector genes. Our data reveal synergistic activation of a multi-layered transcriptional network as the mechanism by which EGFR signaling induces neuronal identity in Glass-expressing cells.
© 2024. The Author(s).
Conflict of interest statement
C.A.M. is a shareholder of 10X Genomics. G.M. is the co-owner of Genetivision Corporation. The other authors declare no competing interests.
Figures



References
MeSH terms
Substances
Grants and funding
- R21EY024826/U.S. Department of Health & Human Services | NIH | Office of Extramural Research, National Institutes of Health (OER)
- R21EY031442/U.S. Department of Health & Human Services | NIH | Office of Extramural Research, National Institutes of Health (OER)
- R21 EY031442/EY/NEI NIH HHS/United States
- R21 EY024826/EY/NEI NIH HHS/United States
- 310030_219348/Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (Swiss National Science Foundation)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
