

MAPT haplotype-associated transcriptomic changes in progressive supranuclear palsy
- PMID: 39154163
- PMCID: PMC11330133
- DOI: 10.1186/s40478-024-01839-3
MAPT haplotype-associated transcriptomic changes in progressive supranuclear palsy
Abstract
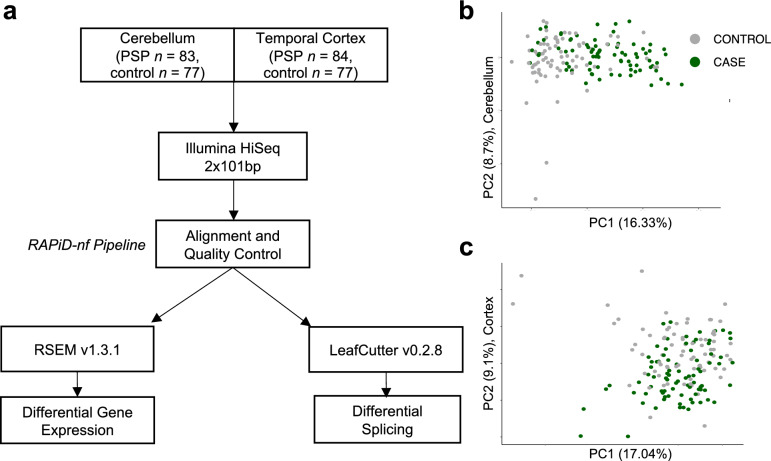
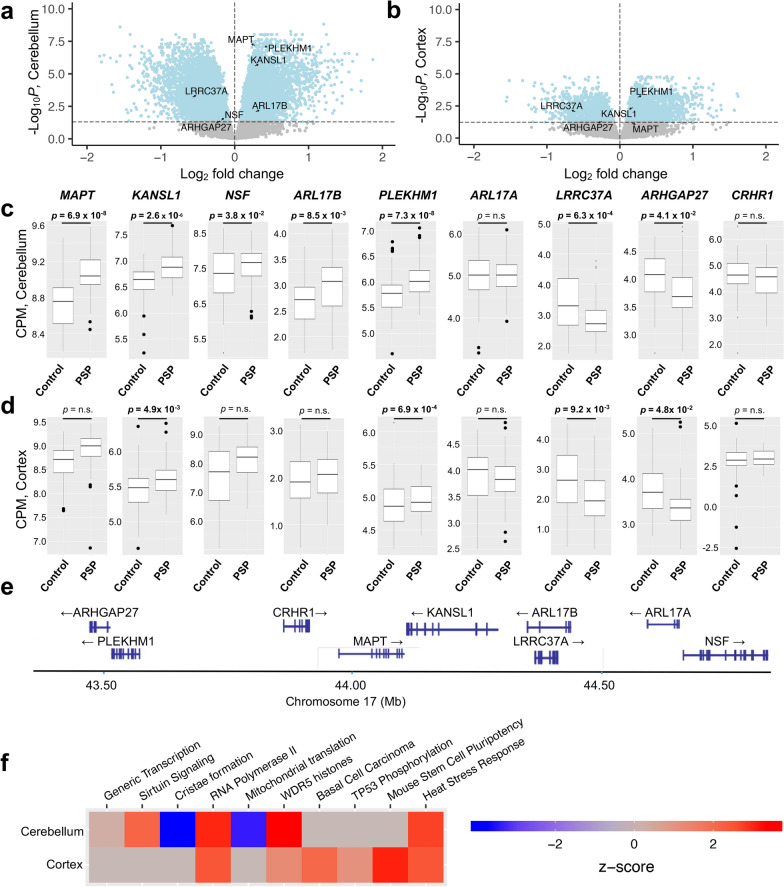
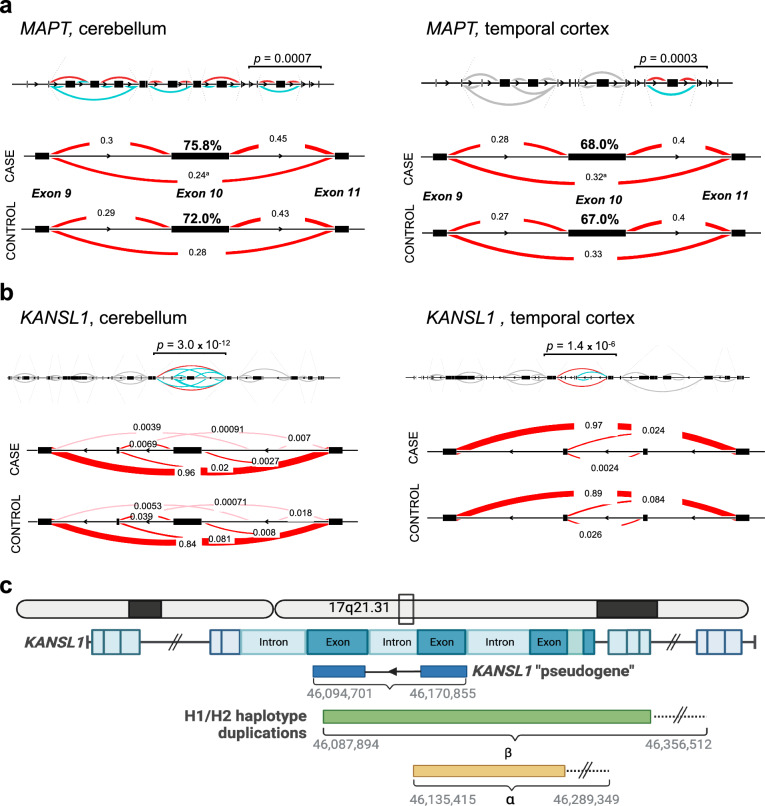
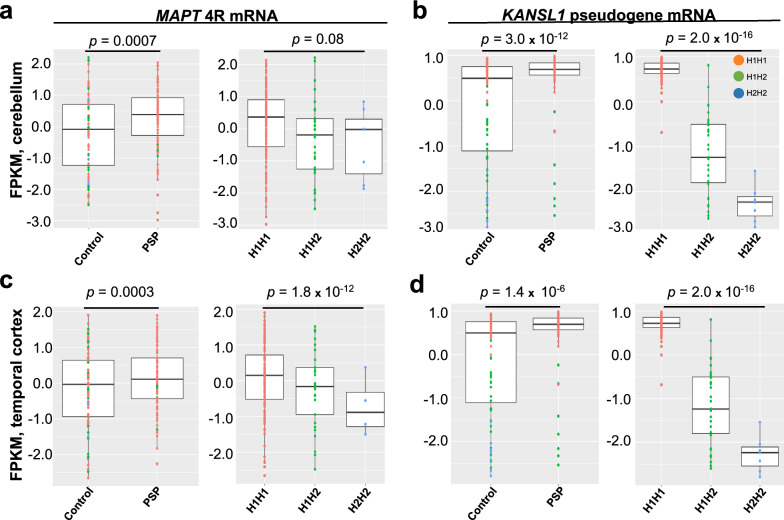
Progressive supranuclear palsy (PSP) is a neurodegenerative movement and cognitive disorder characterized by abnormal accumulation of the microtubule-associated protein tau in the brain. Biochemically, inclusions in PSP are enriched for tau proteoforms with four microtubule-binding domain repeats (4R), an isoform that arises from alternative tau pre-mRNA splicing. While preferential aggregation and reduced degradation of 4R tau protein is thought to play a role in inclusion formation and toxicity, an alternative hypothesis is that altered expression of tau mRNA isoforms plays a causal role. This stems from the observation that PSP is associated with common variation in the tau gene (MAPT) at the 17q21.31 locus which contains low copy number repeats flanking a large recurrent genomic inversion. The complex genomic structural changes at the locus give rise to two dominant haplotypes, termed H1 and H2, that have the potential to markedly influence gene expression. Here, we explored haplotype-dependent differences in gene expression using a bulk RNA-seq dataset derived from human post-mortem brain tissue from PSP (n = 84) and controls (n = 77) using a rigorous computational pipeline, including alternative pre-mRNA splicing. We found 3579 differentially expressed genes in the temporal cortex and 10,011 in the cerebellum. We also found 7214 differential splicing events in the temporal cortex and 18,802 in the cerebellum. In the cerebellum, total tau mRNA levels and the proportion of transcripts encoding 4R tau were significantly increased in PSP compared to controls. In the temporal cortex, the proportion of reads that expressed 4R tau was increased in cases compared to controls. 4R tau mRNA levels were significantly associated with the H1 haplotype in the temporal cortex. Further, we observed a marked haplotype-dependent difference in KANSL1 expression that was strongly associated with H1 in both brain regions. These findings support the hypothesis that sporadic PSP is associated with haplotype-dependent increases in 4R tau mRNA that might play a causal role in this disorder.
Keywords: KANSL1; MAPT haplotype; 17q21.31; Progressive supranuclear palsy; RNA-seq; Tauopathy.
© 2024. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
- R01AG054008/AG/NIA NIH HHS/United States
- T35 AG067578/AG/NIA NIH HHS/United States
- R01NS086736/NS/NINDS NIH HHS/United States
- T35AG067578/AG/NIA NIH HHS/United States
- U54NS115266/NS/NINDS NIH HHS/United States
- R01 AG062348/AG/NIA NIH HHS/United States
- U54 NS115266/NS/NINDS NIH HHS/United States
- R01 NS086736/NS/NINDS NIH HHS/United States
- K01 AG070326/AG/NIA NIH HHS/United States
- P30AG066514/AG/NIA NIH HHS/United States
- P30 AG066514/AG/NIA NIH HHS/United States
- RF1 NS095252/NS/NINDS NIH HHS/United States
- R01 AG054008/AG/NIA NIH HHS/United States
- U19 AG074879/AG/NIA NIH HHS/United States
- RF1NS095252/AG/NIA NIH HHS/United States
- 685-2023-06/CurePSP
- R01AG062348/AG/NIA NIH HHS/United States
- U54NS123743/NS/NINDS NIH HHS/United States
- K01AG070326/AG/NIA NIH HHS/United States
- U54 NS123743/NS/NINDS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
