

Exon 1-targeting miRNA reduces the pathogenic exon 1 HTT protein in Huntington's disease models
- PMID: 39155061
- PMCID: PMC11629698
- DOI: 10.1093/brain/awae266
Exon 1-targeting miRNA reduces the pathogenic exon 1 HTT protein in Huntington's disease models
Abstract
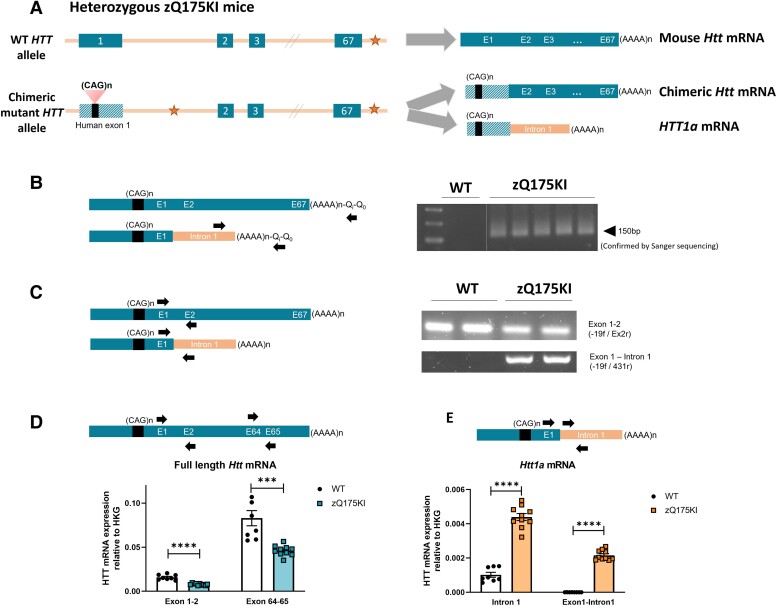
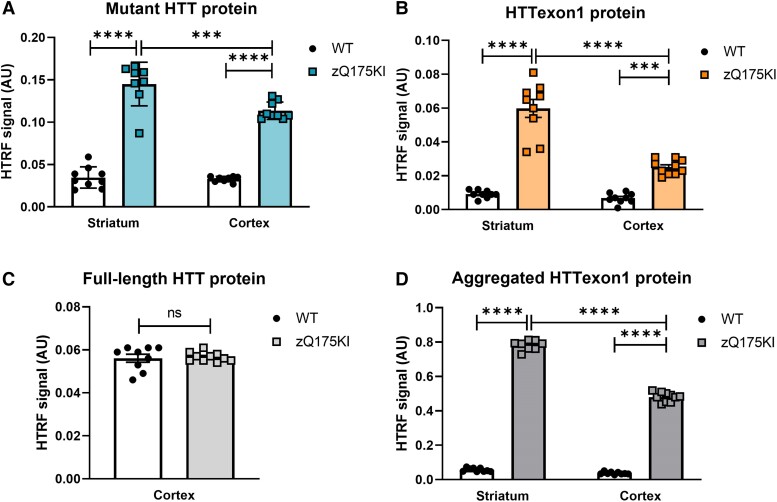
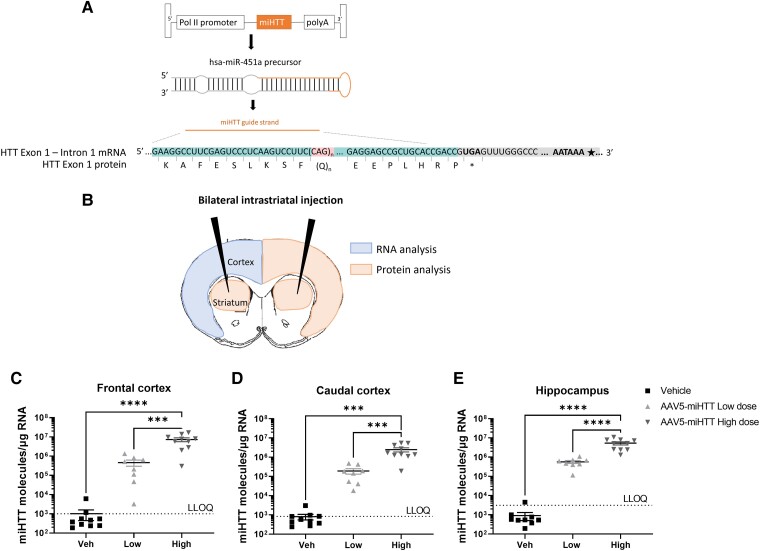
Huntington's disease (HD) is a fatal neurodegenerative disease caused by a trinucleotide repeat expansion in exon 1 of the huntingtin gene (HTT) that results in toxic gain of function and cell death. Despite its monogenic cause, the pathogenesis of HD is highly complex, and increasing evidence indicates that, in addition to the full-length (FL) mutant HTT protein, the expanded exon 1 HTT (HTTexon1) protein that is translated from the HTT1a transcript generated by aberrant splicing is prone to aggregate and might contribute to HD pathology. This finding suggests that reducing the expression of HTT1a might achieve a greater therapeutic benefit than targeting only FL mutant HTT. Conversely, strategies that exclusively target FL HTT might not completely prevent the pathogenesis of HD. We have developed an engineered microRNA targeting the HTT exon 1 sequence (miHTT), delivered via adeno-associated virus serotype 5 (AAV5). The target sequence of miHTT is present in both FL HTT and HTT1a transcripts. Preclinical studies with AAV5-miHTT have demonstrated efficacy in several rodent and large animal models by reducing FL HTT mRNA and protein and rescuing HD-like phenotypes and have been the rationale for phase I/II clinical studies now ongoing in the USA and Europe. In the present study, we evaluated the ability of AAV5-miHTT to reduce the levels of aberrantly spliced HTT1a mRNA and the HTTexon1 protein in the brain of two mouse models of HD (heterozygous zQ175 knock-in mice and humanized Hu128/21 mice). Polyadenylated HTT1a mRNA and HTTexon1 protein were detected in the striatum and cortex of heterozygous zQ175 knock-in mice, but not in wild-type littermate control mice. Intrastriatal administration of AAV5-miHTT resulted in dose-dependent expression of mature miHTT microRNA in cortical brain regions, accompanied by significant lowering of both FL HTT and HTT1a mRNA expression at 2 months postinjection. Mutant HTT and HTTexon1 protein levels were also significantly reduced in the striatum and cortex of heterozygous zQ175 knock-in mice at 2 months after AAV5-miHTT treatment and in humanized Hu128/21 mice 7 months post-treatment. The effects were confirmed in primary Hu128/21 neuronal cultures. These results demonstrate that AAV5-miHTT gene therapy is an effective approach to lower both FL HTT and the pathogenic HTTexon1 levels, which could potentially have an additive therapeutic benefit in comparison to other HTT-targeting modalities.
Keywords: HTT exon 1; AAV5-miHTT; Huntington’s disease; Huntington’s disease mouse models; aberrant splicing; gene therapy.
© The Author(s) 2024. Published by Oxford University Press on behalf of the Guarantors of Brain.
Conflict of interest statement
A.S., A.V. and M.E. are employees of uniQure and may own stock and/or stock options. Filed patent applications pertaining to the results presented in this paper include the following: RNA interference induced
Figures



References
-
- MacDonald ME, Ambrose CM, Duyao MP, et al. . A novel gene containing a trinucleotide repeat that is expanded and unstable on Huntington’s disease chromosomes. Cell. 1993;72:971–983. - PubMed
-
- Tabrizi SJ, Schobel S, Gantman EC, et al. . A biological classification of Huntington’s disease: The integrated staging system. Lancet Neurol. 2022;21:632–644. - PubMed
-
- Stoyas CA, La Spada AR. The CAG–polyglutamine repeat diseases: A clinical, molecular, genetic, and pathophysiologic nosology. In: Geschwind DH, Paulson HL, eds. Handbook of clinical neurology. Vol. 147. Elsevier; 2018:143–170. - PubMed
-
- Takahashi T, Katada S, Onodera O. Polyglutamine diseases: Where does toxicity come from? What is toxicity? Where are we going? J Mol Cell Biol. 2010;2:180–191. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
