

BEAM: A combinatorial recombinase toolbox for binary gene expression and mosaic genetic analysis
- PMID: 39159043
- PMCID: PMC11415793
- DOI: 10.1016/j.celrep.2024.114650
BEAM: A combinatorial recombinase toolbox for binary gene expression and mosaic genetic analysis
Abstract
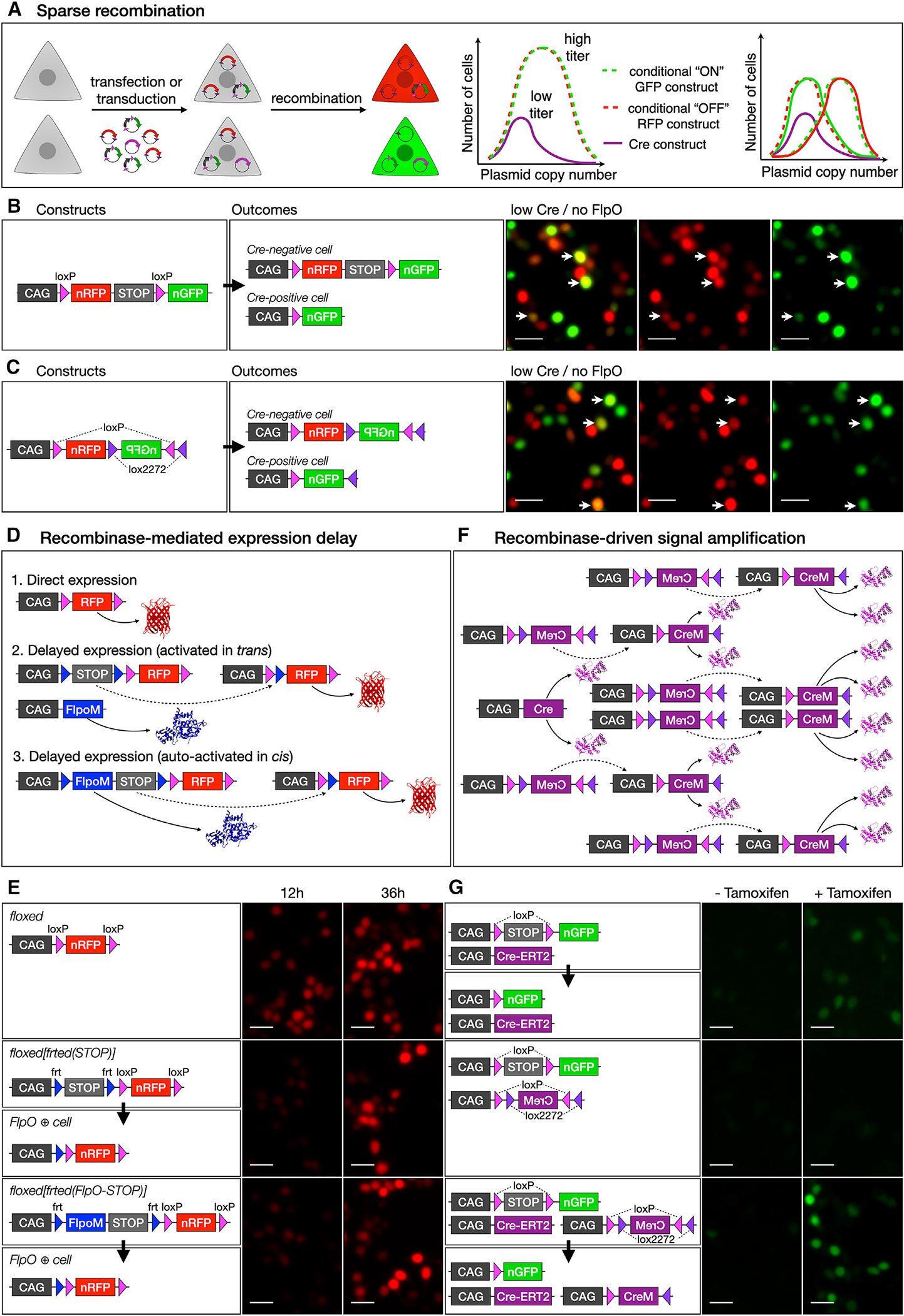
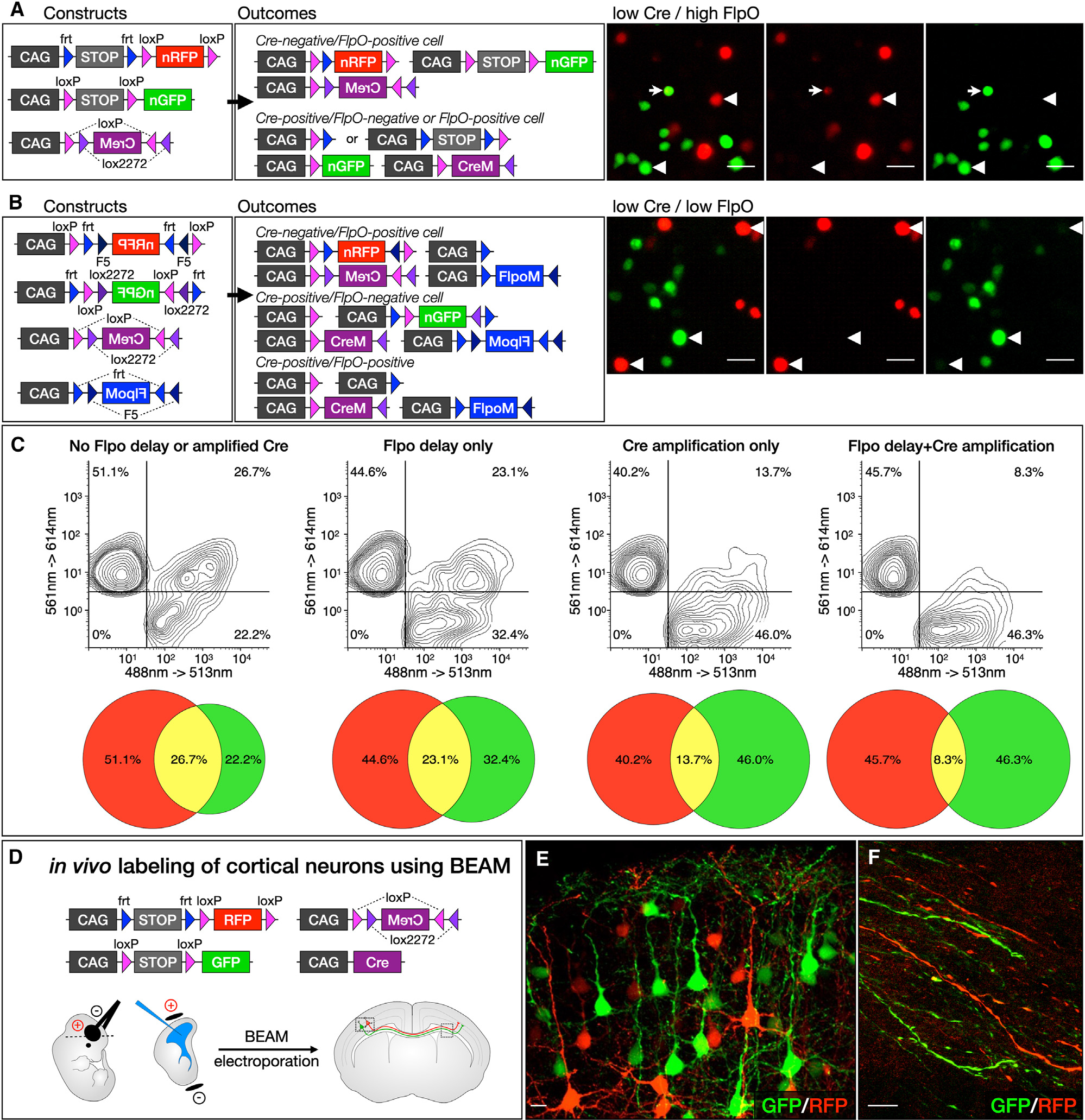
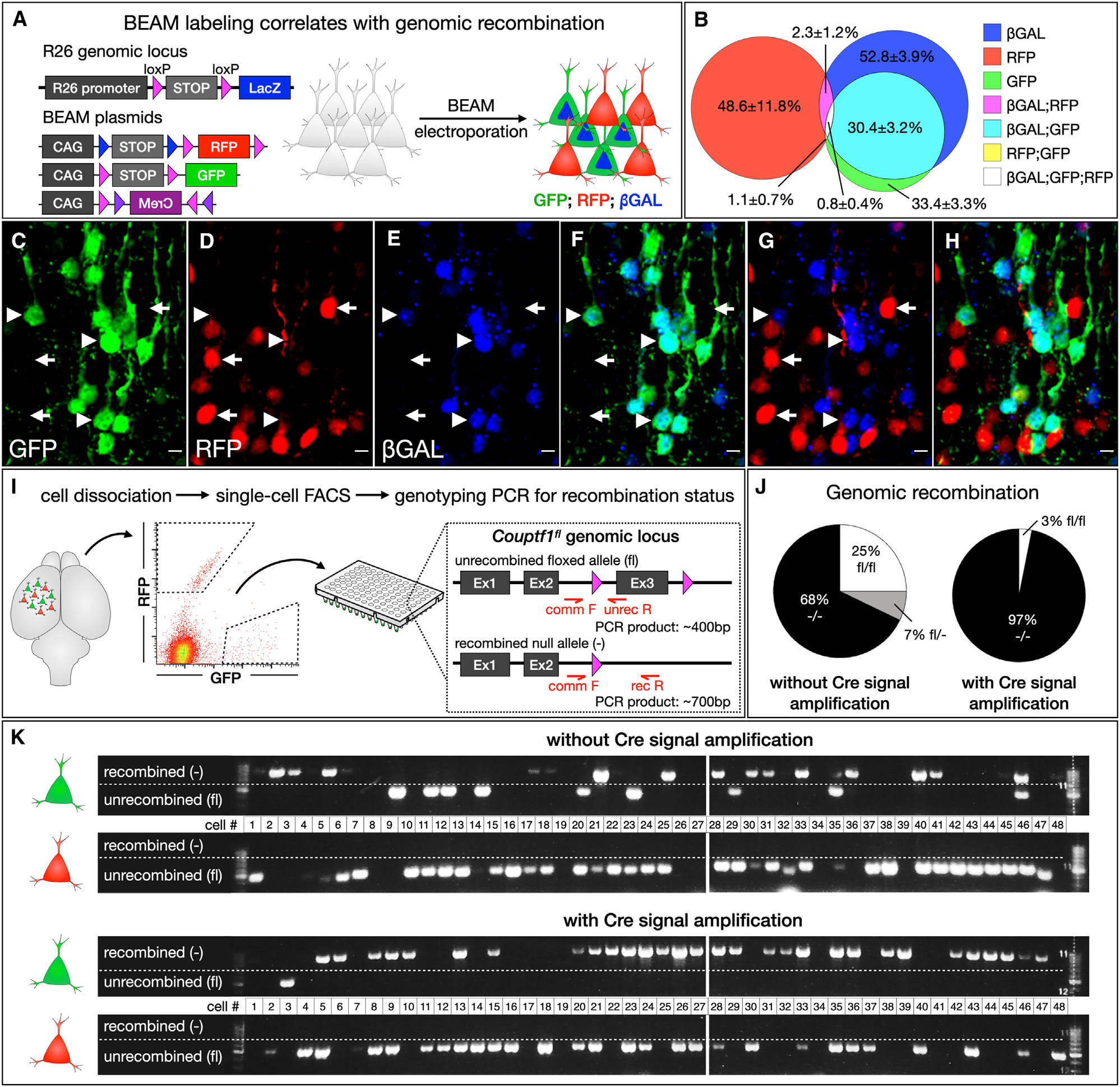
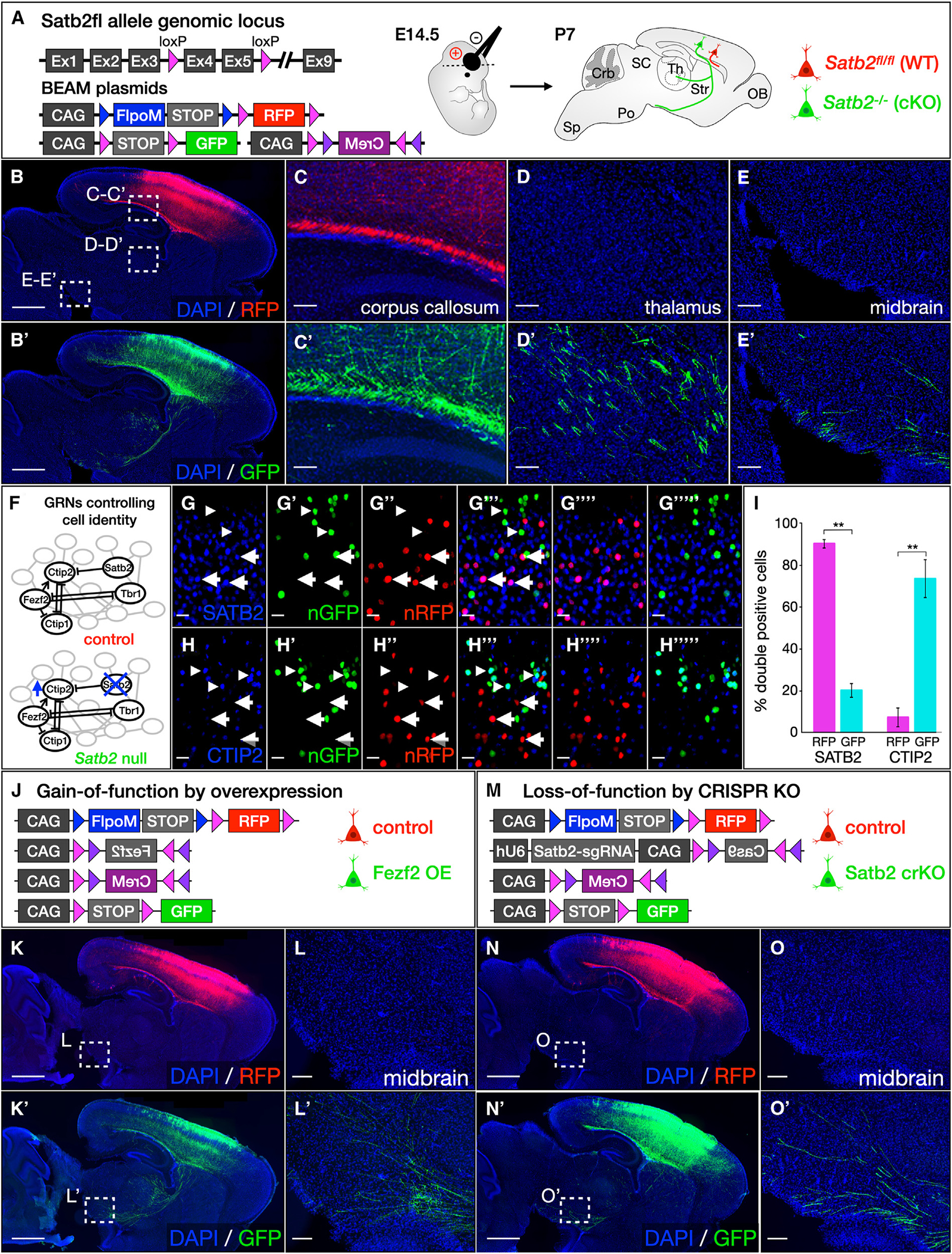
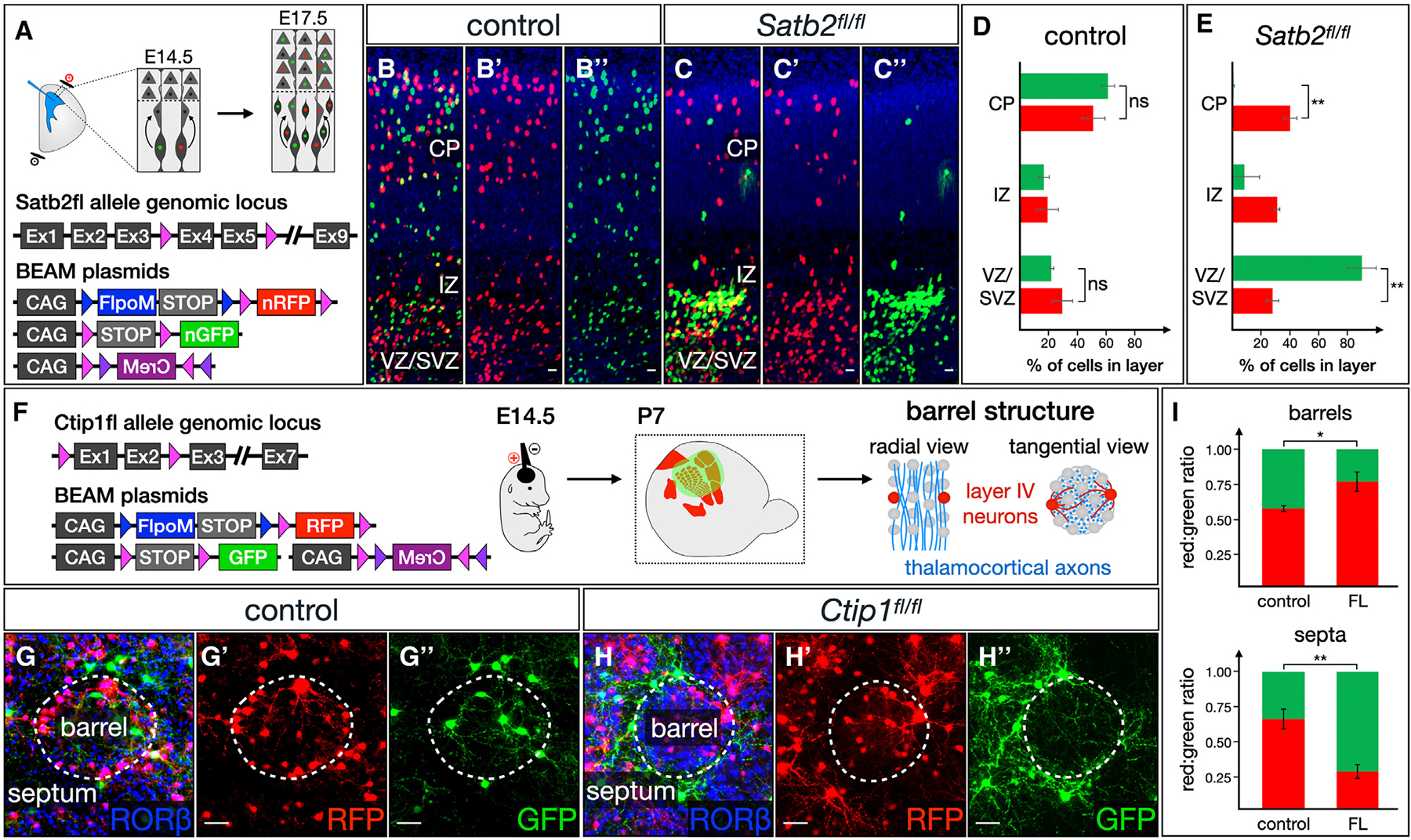
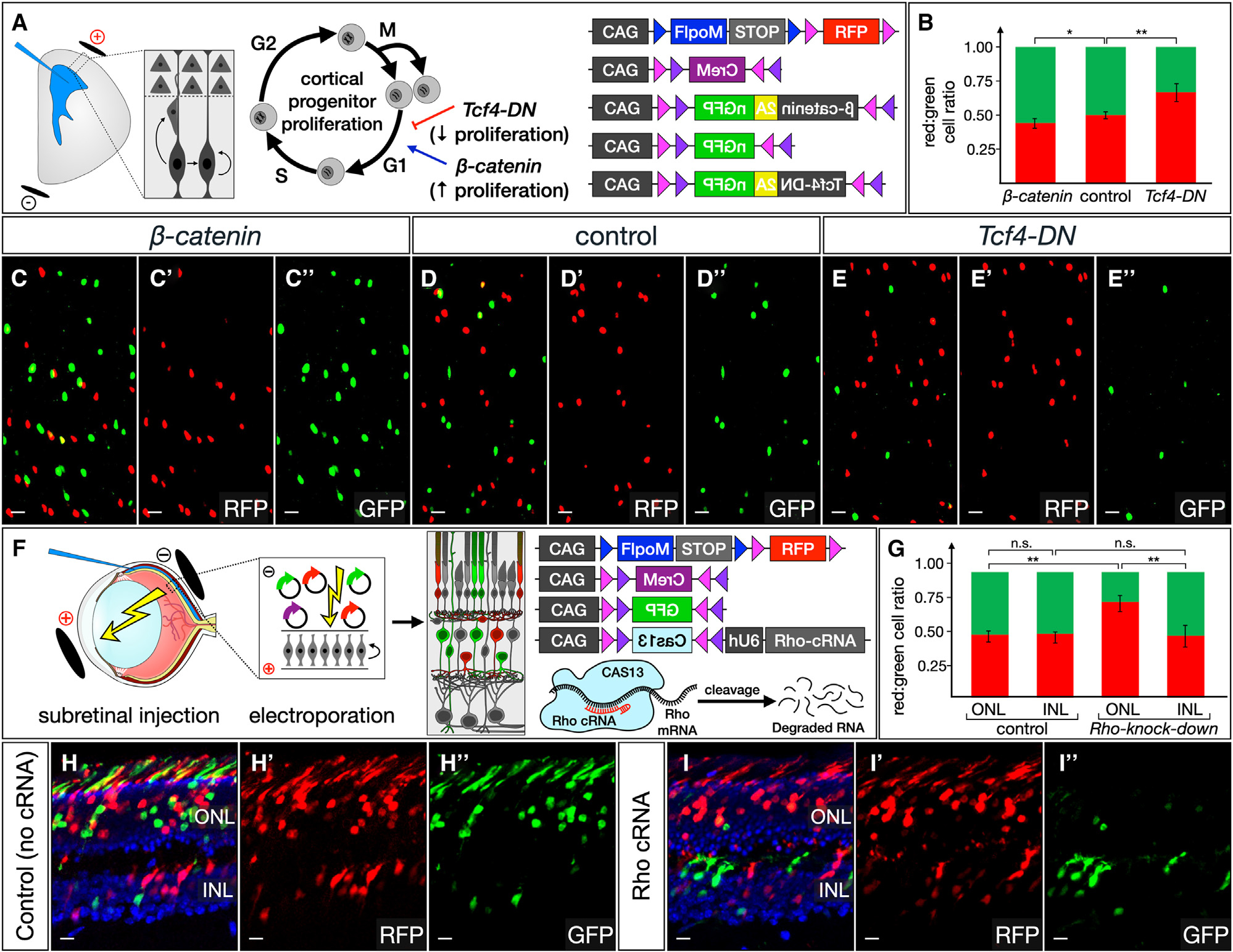
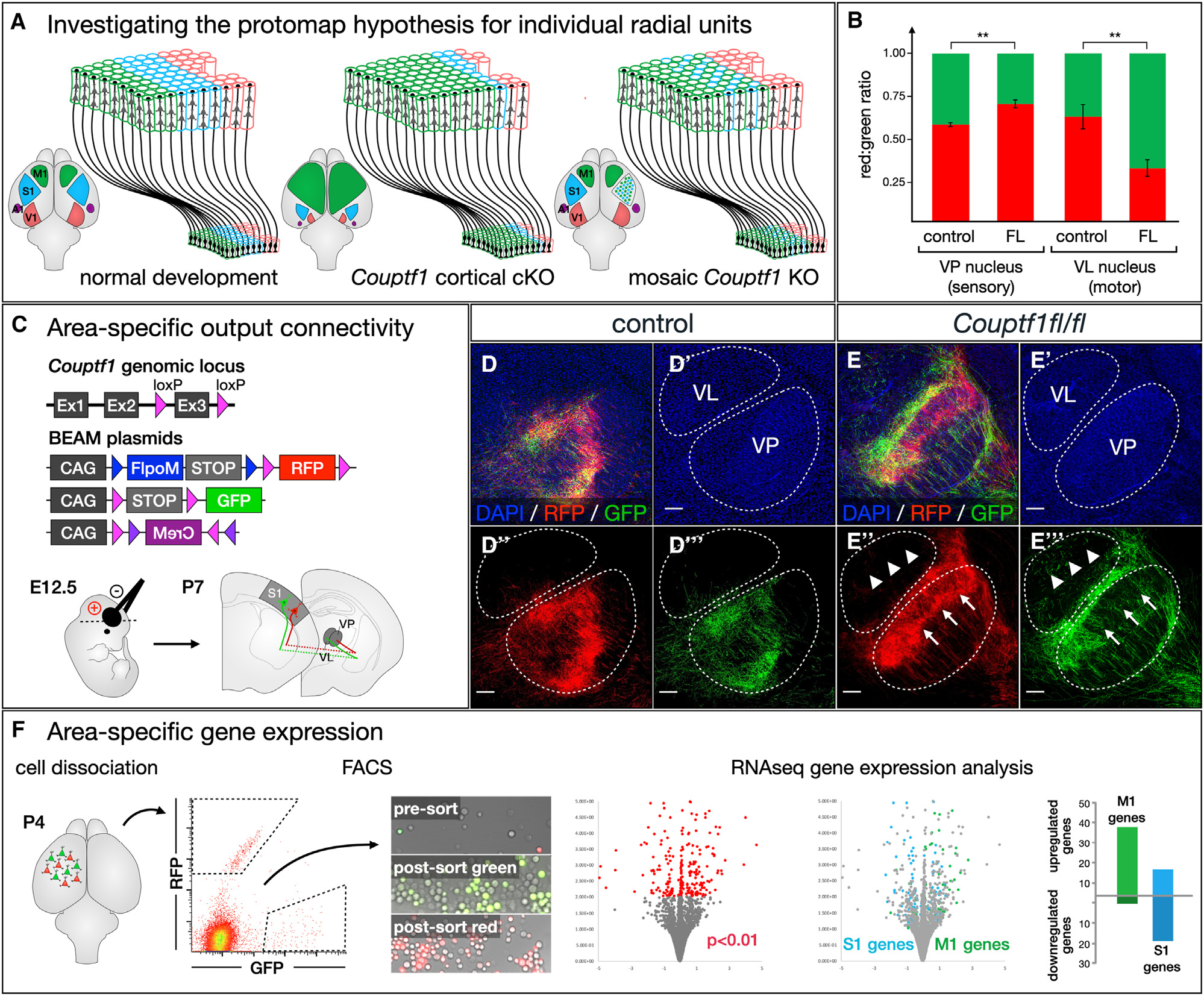
We describe a binary expression aleatory mosaic (BEAM) system, which relies on DNA delivery by transfection or viral transduction along with nested recombinase activity to generate two genetically distinct, non-overlapping populations of cells for comparative analysis. Control cells labeled with red fluorescent protein (RFP) can be directly compared with experimental cells manipulated by genetic gain or loss of function and labeled with GFP. Importantly, BEAM incorporates recombinase-dependent signal amplification and delayed reporter expression to enable sharper delineation of control and experimental cells and to improve reliability relative to existing methods. We applied BEAM to a variety of known phenotypes to illustrate its advantages for identifying temporally or spatially aberrant phenotypes, for revealing changes in cell proliferation or death, and for controlling for procedural variability. In addition, we used BEAM to test the cortical protomap hypothesis at the individual radial unit level, revealing that area identity is cell autonomously specified in adjacent radial units.
Keywords: CP: Cell biology; Cre; Flp; genetic analysis; mosaicism; recombinase.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Update of
-
BEAM: a combinatorial recombinase toolbox for binary gene expression and mosaic analysis.bioRxiv [Preprint]. 2023 Feb 16:2023.02.16.528875. doi: 10.1101/2023.02.16.528875. bioRxiv. 2023. Update in: Cell Rep. 2024 Aug 27;43(8):114650. doi: 10.1016/j.celrep.2024.114650. PMID: 36824714 Free PMC article. Updated. Preprint.
References
-
- Villalba A, Amberg N, and Hippenmeyer S (2023). Interplay of cell-autonomous gene function and tissue-wide mechanisms regulating radial glial progenitor lineage progression. Neocortical Neurogenesis in Development and Evolution, 169–191. 10.1002/9781119860914. - DOI
-
- Zong H, Espinosa JS, Su HH, Muzumdar MD, and Luo L (2005). Mosaic analysis with double markers in mice. Cell 121, 479–492. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
