

Multiple guidance mechanisms control axon growth to generate precise T-shaped bifurcation during dorsal funiculus development in the spinal cord
- PMID: 39159057
- PMCID: PMC11333043
- DOI: 10.7554/eLife.94109
Multiple guidance mechanisms control axon growth to generate precise T-shaped bifurcation during dorsal funiculus development in the spinal cord
Abstract
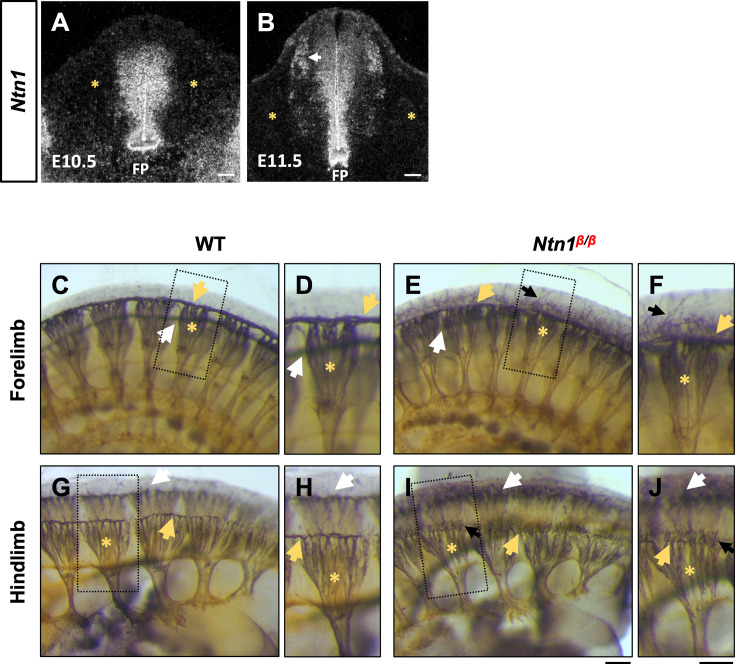
The dorsal funiculus in the spinal cord relays somatosensory information to the brain. It is made of T-shaped bifurcation of dorsal root ganglion (DRG) sensory axons. Our previous study has shown that Slit signaling is required for proper guidance during bifurcation, but loss of Slit does not affect all DRG axons. Here, we examined the role of the extracellular molecule Netrin-1 (Ntn1). Using wholemount staining with tissue clearing, we showed that mice lacking Ntn1 had axons escaping from the dorsal funiculus at the time of bifurcation. Genetic labeling confirmed that these misprojecting axons come from DRG neurons. Single axon analysis showed that loss of Ntn1 did not affect bifurcation but rather altered turning angles. To distinguish their guidance functions, we examined mice with triple deletion of Ntn1, Slit1, and Slit2 and found a completely disorganized dorsal funiculus. Comparing mice with different genotypes using immunolabeling and single axon tracing revealed additive guidance errors, demonstrating the independent roles of Ntn1 and Slit. Moreover, the same defects were observed in embryos lacking their cognate receptors. These in vivo studies thus demonstrate the presence of multi-factorial guidance mechanisms that ensure proper formation of a common branched axonal structure during spinal cord development.
Keywords: axon bifurcation; developmental biology; dorsal funiculus; mouse; neuroscience; spinal cord.
© 2024, Curran et al.
Conflict of interest statement
BC, KN, AY, LG, AJ, MT, LM No competing interests declared
Figures








Update of
-
Multiple Guidance Mechanisms Control Axon Growth to Generate Precise T-shaped Bifurcation during Dorsal Funiculus Development in the Spinal Cord.bioRxiv [Preprint]. 2024 Apr 15:2023.11.17.567638. doi: 10.1101/2023.11.17.567638. bioRxiv. 2024. Update in: Elife. 2024 Aug 19;13:RP94109. doi: 10.7554/eLife.94109. PMID: 38014092 Free PMC article. Updated. Preprint.
References
-
- Bin JM, Han D, Lai Wing Sun K, Croteau L-P, Dumontier E, Cloutier J-F, Kania A, Kennedy TE. Complete loss of netrin-1 results in embryonic lethality and severe axon guidance defects without increased neural cell death. Cell Reports. 2015;12:1099–1106. doi: 10.1016/j.celrep.2015.07.028. - DOI - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
