

ISGylation enhances dsRNA-induced interferon response and NFκB signaling in fallopian tube epithelial cells
- PMID: 39159817
- PMCID: PMC11418117
- DOI: 10.1016/j.jbc.2024.107686
ISGylation enhances dsRNA-induced interferon response and NFκB signaling in fallopian tube epithelial cells
Abstract
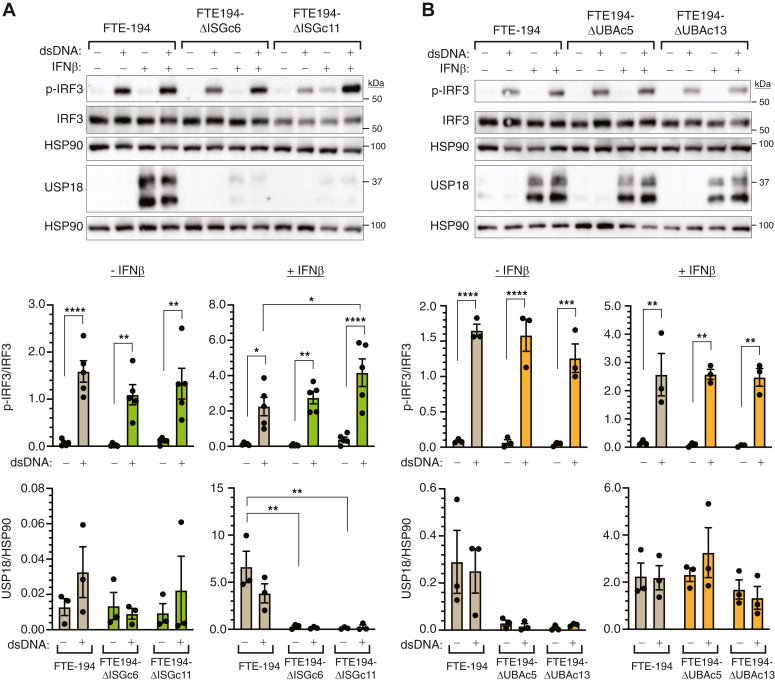
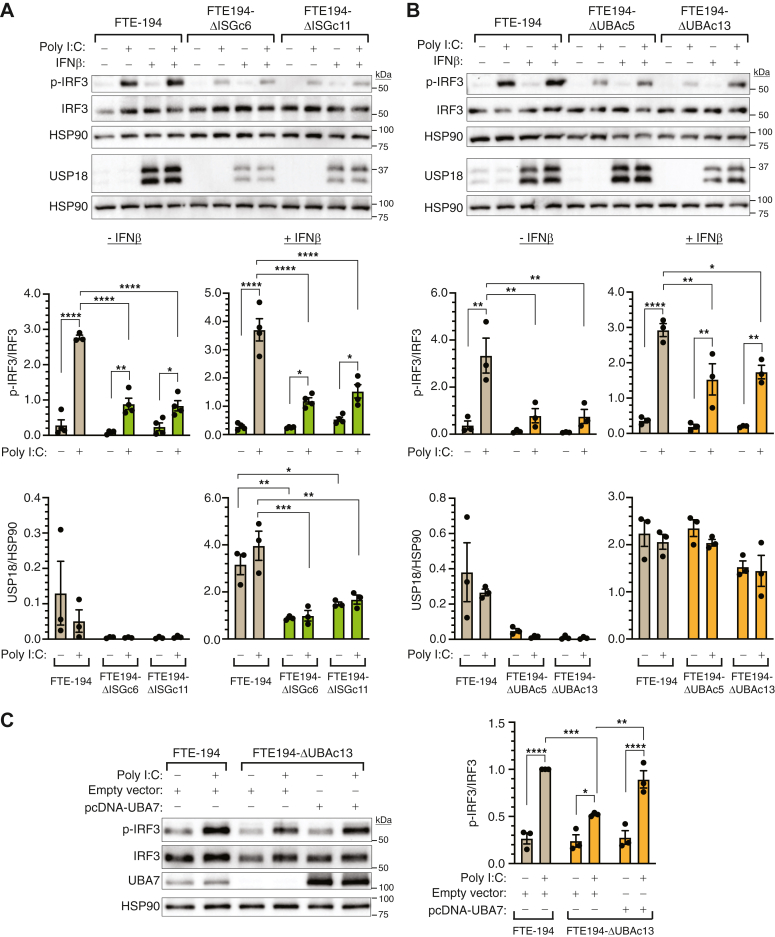
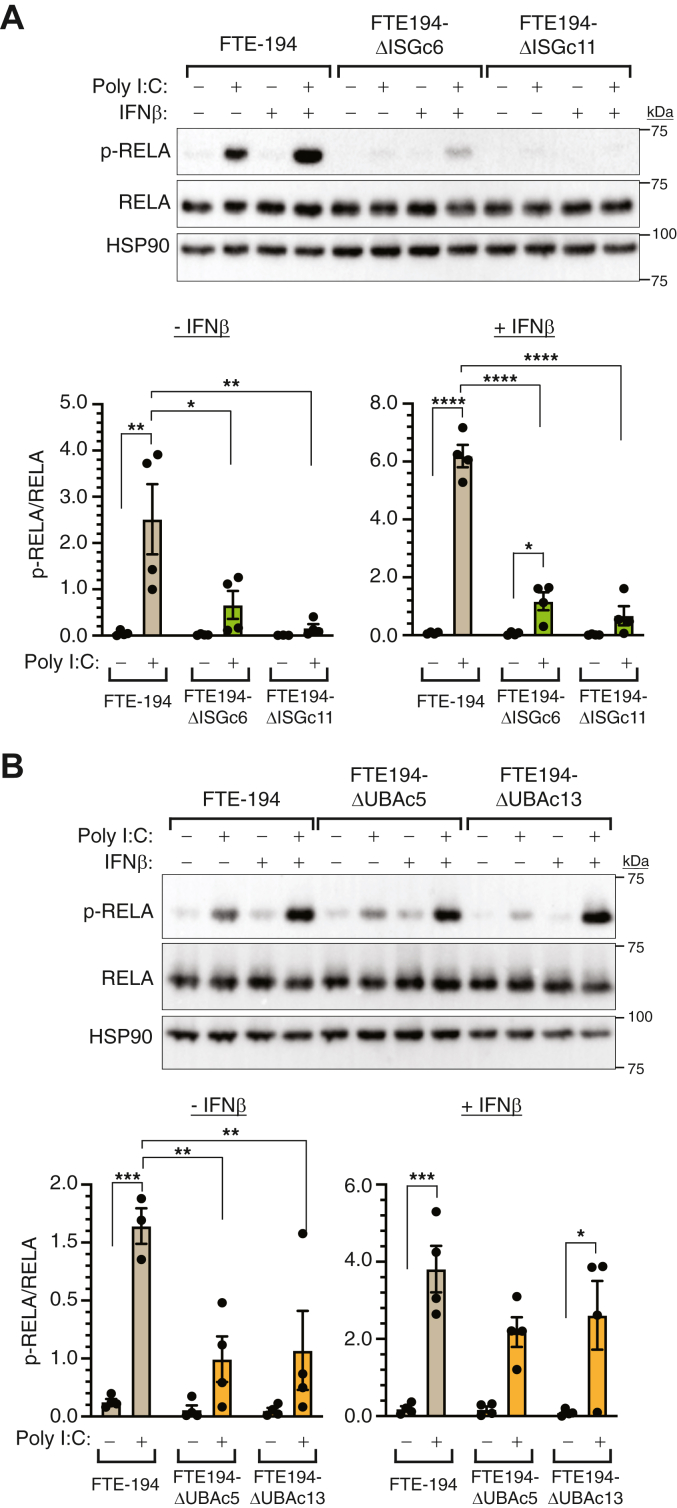
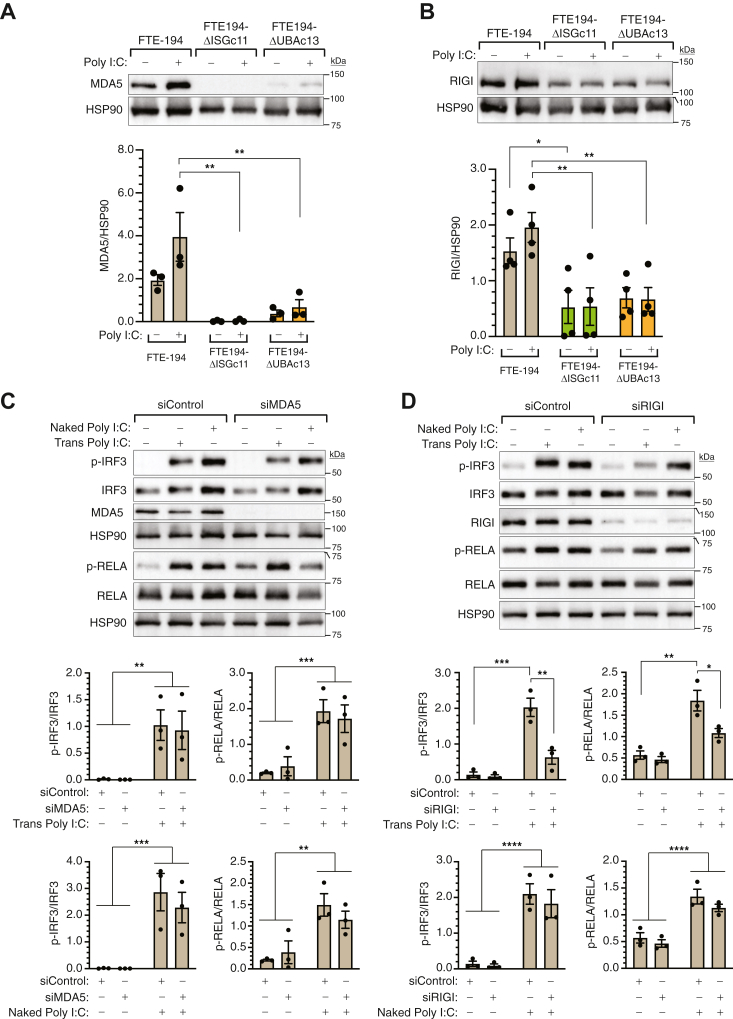
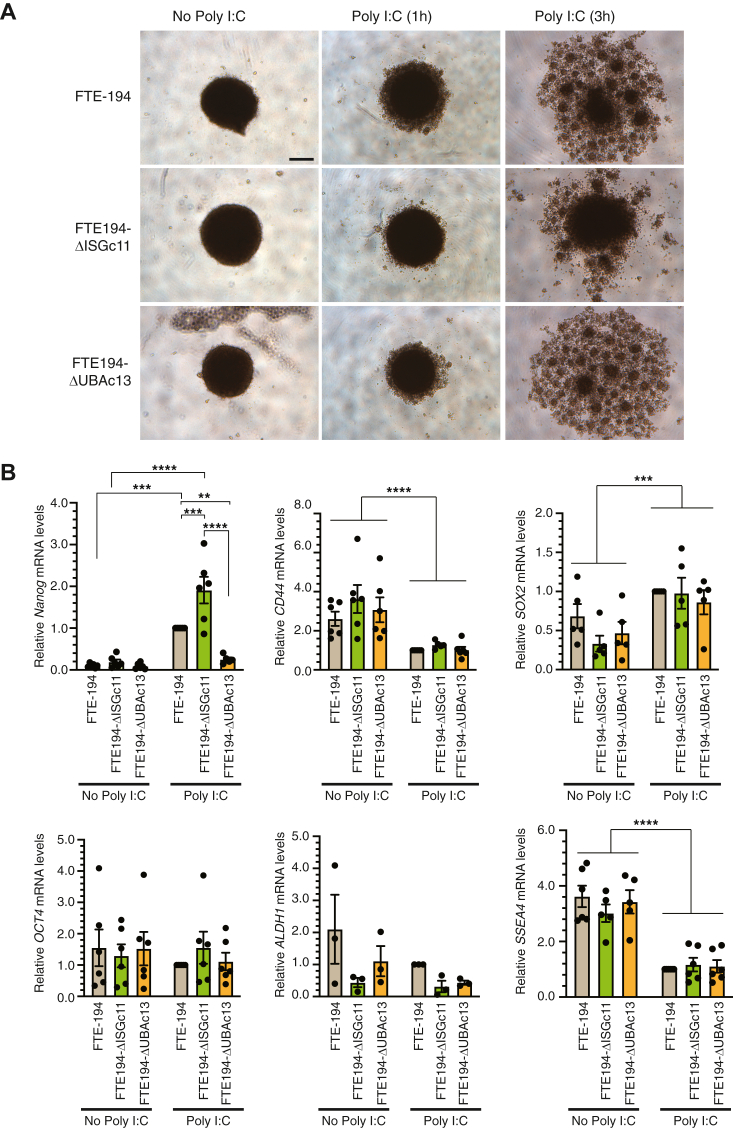
Heritable mutations in BRCA1 associate with increased risk of high-grade serous tubo-ovarian cancer. Nongenetic risk factors associated with this cancer, which arises from fallopian tube epithelial (FTE) cells, suggests a role for repetitive ovulation wherein FTE cells are exposed to inflammatory signaling molecules within follicular fluid. We previously reported increased NFκB and EGFR signaling in BRCA1-deficient primary FTE cells, with follicular fluid exposure further increasing abundance of interferon-stimulated gene (ISG) transcripts, including the ubiquitin-like protein ISG15 and other ISGylation pathway members. Both NFκB and type I interferon signaling are upregulated by stimulation of cGAS-STING or MDA5 and RIGI pattern recognition receptors. Since some pattern recognition receptors and their signal transduction pathway members are ISGylated, we tested the impact of ISG15 and ISGylation on interferon regulatory factor 3 (IRF3) and NFκB signaling through cGAS-STING or RIGI and MDA5 activation. Expression of ISG15 or UBA7, the E1-like ISG15-activating enzyme, in immortalized FTE cells was disrupted by CRISPR gene editing. Activation of IRF3 by RIGI or MDA5 but not cGAS-STING was attenuated by loss of either ISG15 or UBA7 and this was reflected by a similar effect on NFκB activation and downstream targets. Loss of ISGylation decreased levels of both MDA5 and RIGI, with knockdown of RIGI but not MDA5, decreasing IRF3 and NFκB activation in parental cells. These finding indicate that ISGylation enhances the ability of dsRNA to activate cytokine release and proinflammatory signaling. Further work to explore ISGylation as a target for prevention of high-grade serous tubo-ovarian cancer in BRCA1 mutation carriers is warranted.
Keywords: CRISPR/cas; ISGylation; NF-kappa B; RIGI-like receptor; dsRNA; fallopian tube cells; inflammation; innate immunity; interferon; ovarian cancer.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures





References
-
- Kuchenbaecker K.B., Hopper J.L., Barnes D.R., Phillips K.A., Mooij T.M., Roos-Blom M.J., et al. Risks of breast, ovarian, and contralateral breast cancer for BRCA1 and BRCA2 mutation carriers. JAMA. 2017;317:2402–2416. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
