

An extrinsic motor directs chromatin loop formation by cohesin
- PMID: 39160275
- PMCID: PMC11445435
- DOI: 10.1038/s44318-024-00202-5
An extrinsic motor directs chromatin loop formation by cohesin
Erratum in
-
Author Correction: An extrinsic motor directs chromatin loop formation by cohesin.EMBO J. 2025 Mar;44(5):1563-1568. doi: 10.1038/s44318-024-00341-9. EMBO J. 2025. PMID: 39890887 Free PMC article.
Abstract
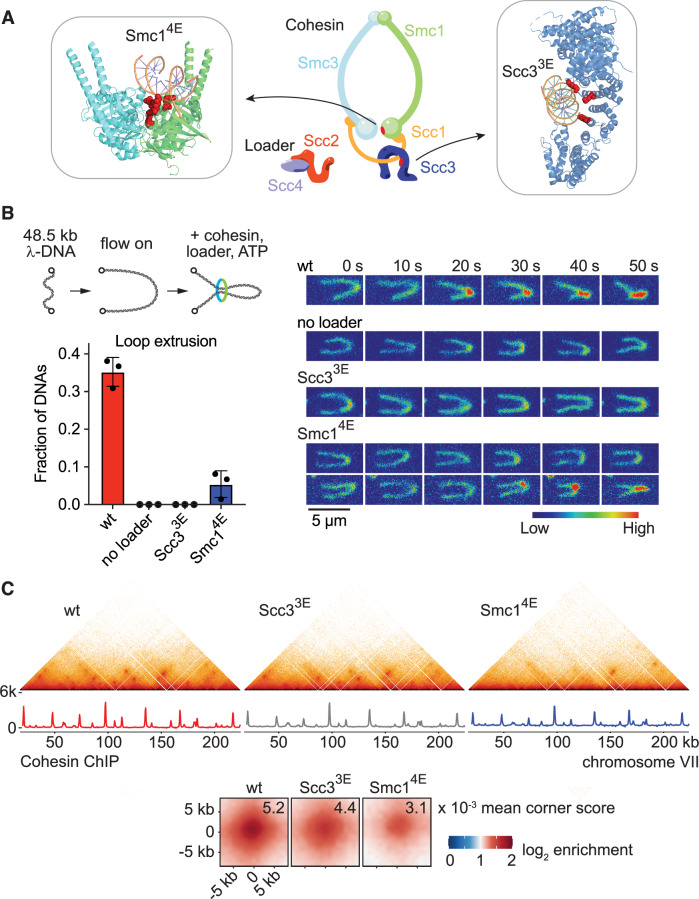
The ring-shaped cohesin complex topologically entraps two DNA molecules to establish sister chromatid cohesion. Cohesin also shapes the interphase chromatin landscape with wide-ranging implications for gene regulation, and cohesin is thought to achieve this by actively extruding DNA loops without topologically entrapping DNA. The 'loop extrusion' hypothesis finds motivation from in vitro observations-whether this process underlies in vivo chromatin loop formation remains untested. Here, using the budding yeast S. cerevisiae, we generate cohesin variants that have lost their ability to extrude DNA loops but retain their ability to topologically entrap DNA. Analysis of these variants suggests that in vivo chromatin loops form independently of loop extrusion. Instead, we find that transcription promotes loop formation, and acts as an extrinsic motor that expands these loops and defines their ultimate positions. Our results necessitate a re-evaluation of the loop extrusion hypothesis. We propose that cohesin, akin to sister chromatid cohesion establishment at replication forks, forms chromatin loops by DNA-DNA capture at places of transcription, thus unifying cohesin's two roles in chromosome segregation and interphase genome organisation.
Keywords: Cohesin; Loop Capture; Loop Extrusion; SMC Complexes; Transcription.
© 2024. The Author(s).
Figures










References
-
- Baejen C et al (2017) Genome-wide analysis of RNA Polymerase II termination at protein-coding genes. Mol Cell 66:38–49 - PubMed
-
- Barth R, Davidson IF, van der Torre J, Taschner M, Gruber S, Peters J-M, Dekker C (2023) SMC motor proteins extrude DNA asymmetrically and contain a direction switch. Preprint at bioRxiv 10.1101/2023.12.21.572892
-
- Bastié N, Chapard C, Dauban L, Gadal O, Beckouët F, Koszul R (2022) Smc3 acetylation, Pds5 and Scc2 control the translocase activity that establishes cohesin-dependent chromatin loops. Nat Struct Mol Biol 29:575–585 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
