

DNA Methylation-Mediated GPX4 Transcriptional Repression and Osteoblast Ferroptosis Promote Titanium Particle-Induced Osteolysis
- PMID: 39161535
- PMCID: PMC11331012
- DOI: 10.34133/research.0457
DNA Methylation-Mediated GPX4 Transcriptional Repression and Osteoblast Ferroptosis Promote Titanium Particle-Induced Osteolysis
Abstract
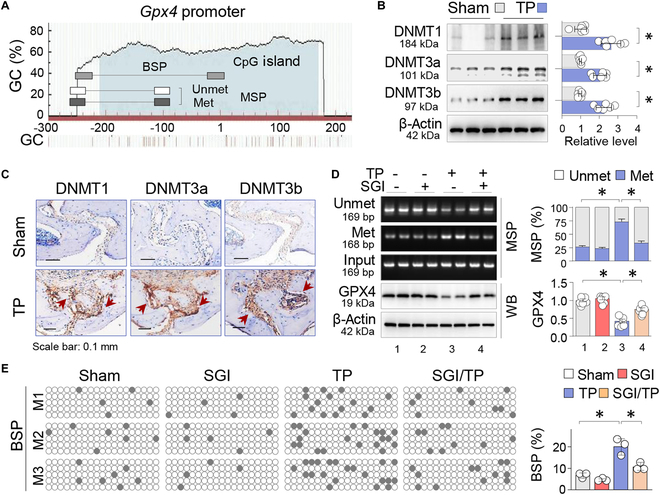
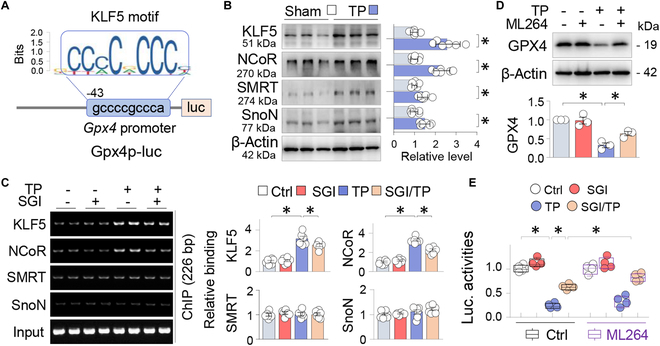
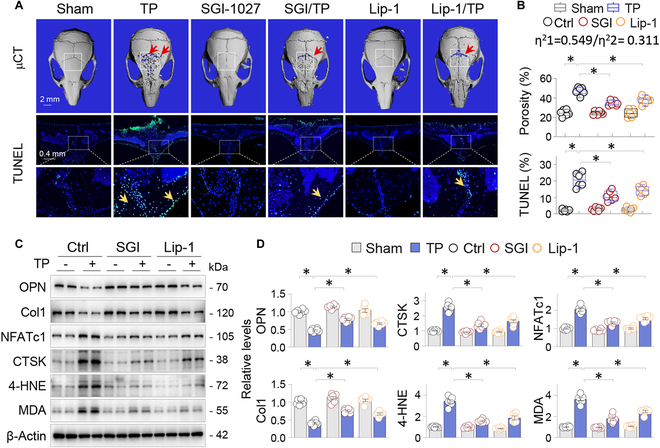
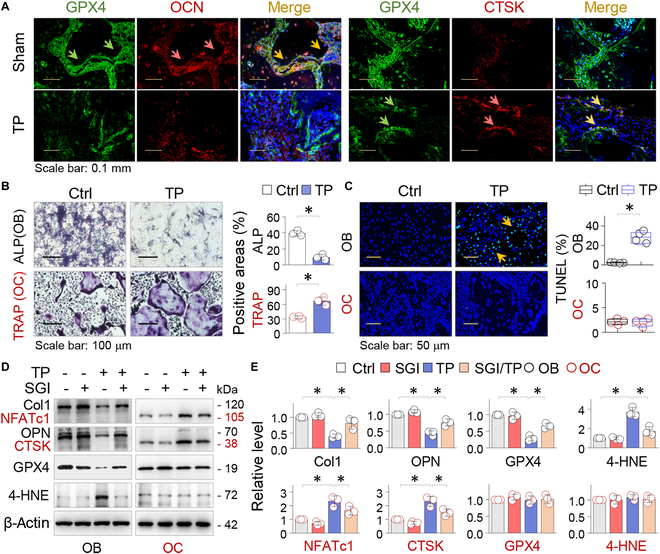
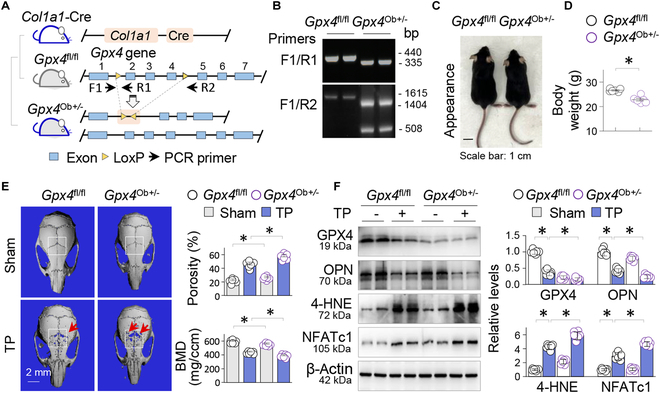
Metal wear particles generated by the movement of joint prostheses inevitably lead to aseptic osteolytic damage and ultimately prosthesis loosening, which are aggravated by various types of regulated cell death of bone. Nevertheless, the exact cellular nature and regulatory network underlying osteoferroptosis are poorly understood. Here, we report that titanium particles (TP) induced severe peri-implant osteolysis and ferroptotic changes with concomitant transcriptional repression of a key anti-ferroptosis factor, GPX4, in a mouse model of calvarial osteolysis. GPX4 repression was accompanied by an increase in DNA methyltransferases (DNMTs) 1/3a/3b and hypermethylation of the Gpx4 promoter, which were partly mediated by the transcriptional regulator/co-repressor KLF5 and NCoR. Conversely, treatment with SGI-1027, a DNMT-specific inhibitor, resulted in marked reversal of Gpx4 promoter hypermethylation and GPX4 repression, as well as improvement in ferroptotic osteolysis to a similar extent as with a ferroptosis inhibitor, liproxstatin-1. This suggests that epigenetic GPX4 repression and ferroptosis caused by the increase of DNMT1/3a/3b have a causal influence on TP-induced osteolysis. In cultured primary osteoblasts and osteoclasts, GPX4 repression and ferroptotic changes were observed primarily in osteoblasts that were alleviated by SGI-1027 in a GPX4 inactivation-sensitive manner. Furthermore, we developed a mouse strain with Gpx4 haplodeficiency in osteoblasts (Gpx4 Ob+/-) that exhibited worsened ferroptotic osteolysis in control and TP-treated calvaria and largely abolished the anti-ferroptosis and osteoprotective effects of SGI-1027. Taken together, our results demonstrate that DNMT1/3a/3b elevation, resulting GPX4 repression, and osteoblastic ferroptosis form a critical epigenetic pathway that significantly contributes to TP-induced osteolysis, and that targeting DNMT aberration and the associated osteoferroptosis could be a potential strategy to prevent or slow down prosthesis-related osteolytic complications.
Copyright © 2024 Jian Dong et al.
Conflict of interest statement
Competing interests: The authors declare that they have no competing interests.
Figures



References
-
- Purdue PE, Koulouvaris P, Potter HG, Nestor BJ, Sculco TP. The cellular and molecular biology of periprosthetic osteolysis. Clin Orthop Relat Res. 2007;454:251–261. - PubMed
-
- Wauquier F, Leotoing L, Coxam V, Guicheux J, Wittrant Y. Oxidative stress in bone remodelling and disease. Trends Mol Med. 2009;15(10):468–477. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
