

Clinical immunity to malaria involves epigenetic reprogramming of innate immune cells
- PMID: 39161730
- PMCID: PMC11331423
- DOI: 10.1093/pnasnexus/pgae325
Clinical immunity to malaria involves epigenetic reprogramming of innate immune cells
Abstract
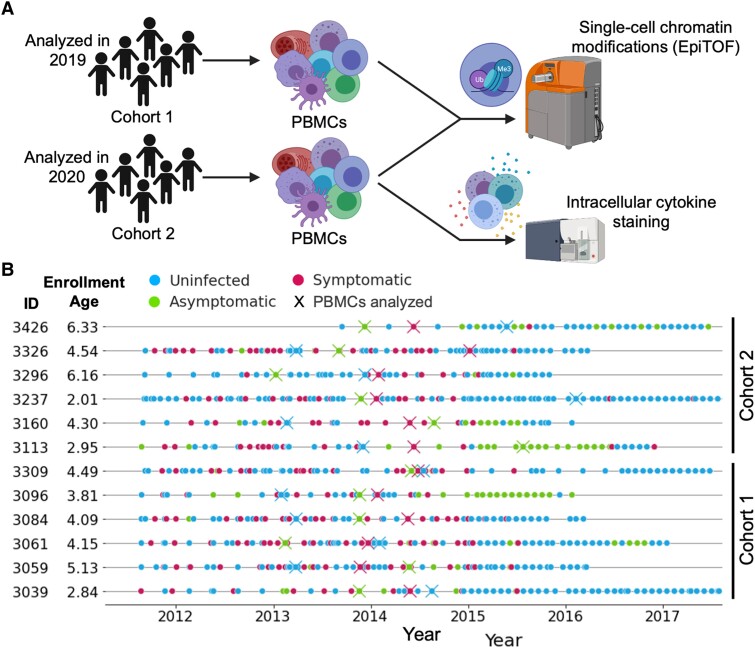
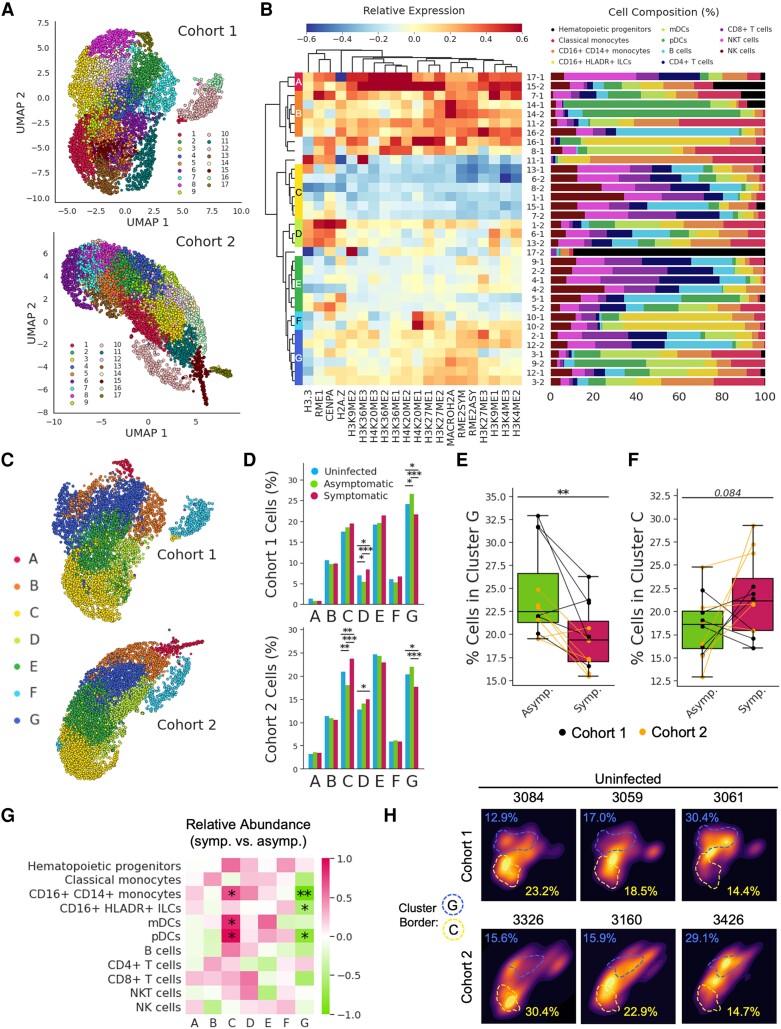
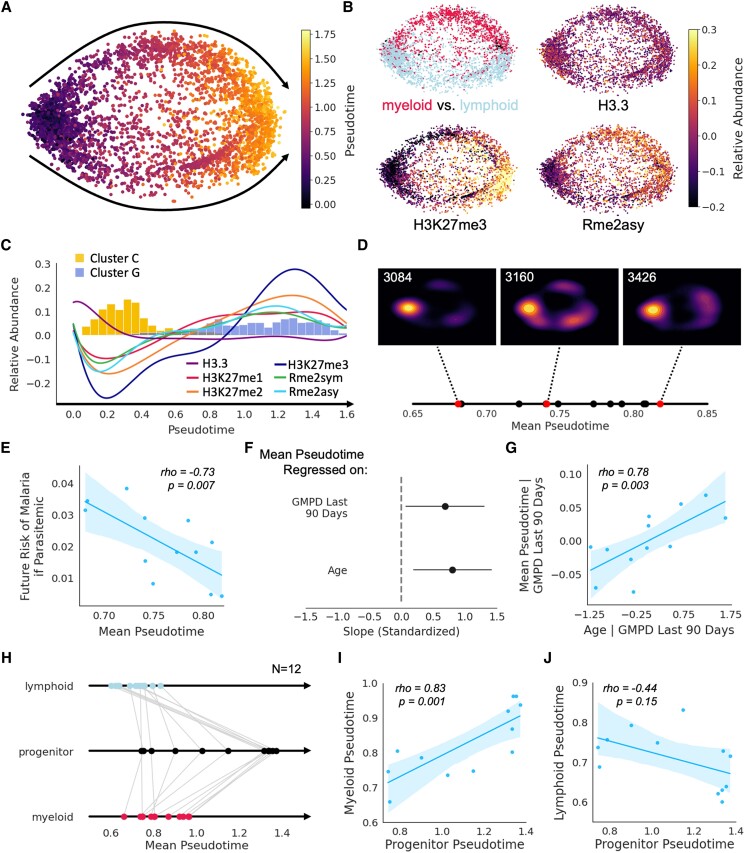
The regulation of inflammation is a critical aspect of disease tolerance and naturally acquired clinical immunity to malaria. Here, we demonstrate using RNA sequencing and epigenetic landscape profiling by cytometry by time-of-flight, that the regulation of inflammatory pathways during asymptomatic parasitemia occurs downstream of pathogen sensing-at the epigenetic level. The abundance of certain epigenetic markers (methylation of H3K27 and dimethylation of arginine residues) and decreased prevalence of histone variant H3.3 correlated with suppressed cytokine responses among monocytes of Ugandan children. Such an epigenetic signature was observed across diverse immune cell populations and not only characterized active asymptomatic parasitemia but also correlated with future long-term disease tolerance and clinical immunity when observed in uninfected children. Pseudotime analyses revealed a potential trajectory of epigenetic change that correlated with a child's age and recent parasite exposure and paralleled the acquisition of clinical immunity. Thus, our data support a model whereby exposure to Plasmodium falciparum induces epigenetic changes that regulate excessive inflammation and contribute to naturally acquire clinical immunity to malaria.
© The Author(s) 2024. Published by Oxford University Press on behalf of National Academy of Sciences.
Figures




References
-
- World Health Organization . 2022. World malaria report 2022. Geneva: World Health Organization.
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
