

Prevotella timonensis degrades the vaginal epithelial glycocalyx through high fucosidase and sialidase activities
- PMID: 39162399
- PMCID: PMC11389373
- DOI: 10.1128/mbio.00691-24
Prevotella timonensis degrades the vaginal epithelial glycocalyx through high fucosidase and sialidase activities
Abstract
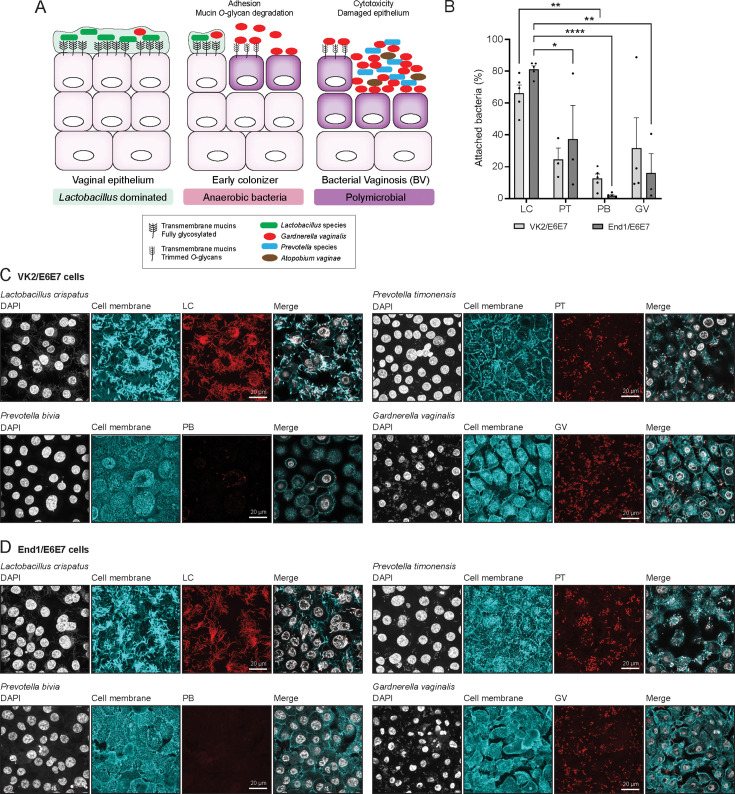
Bacterial vaginosis (BV) is a polymicrobial infection of the female reproductive tract. BV is characterized by replacement of health-associated Lactobacillus species by diverse anerobic bacteria, including the well-known Gardnerella vaginalis. Prevotella timonensis, and Prevotella bivia are anerobes that are found in a significant number of BV patients, but their contributions to the disease process remain to be determined. Defining characteristics of anerobic overgrowth in BV are adherence to the mucosal surface and the increased activity of mucin-degrading enzymes such as sialidases in vaginal secretions. We demonstrate that P. timonensis, but not P. bivia, strongly adheres to vaginal and endocervical cells to a similar level as G. vaginalis but did not elicit a comparable proinflammatory epithelial response. The P. timonensis genome uniquely encodes a large set of mucus-degrading enzymes, including four putative fucosidases and two putative sialidases, PtNanH1 and PtNanH2. Enzyme assays demonstrated that fucosidase and sialidase activities in P. timonensis cell-bound and secreted fractions were significantly higher than for other vaginal anerobes. In infection assays, P. timonensis efficiently removed fucose and α2,3- and α2,6-linked sialic acid moieties from the epithelial glycocalyx. Recombinantly expressed P. timonensis NanH1 and NanH2 cleaved α2,3 and α2,6-linked sialic acids from the epithelial surface, and sialic acid removal by P. timonensis could be blocked using inhibitors. This study demonstrates that P. timonensis has distinct virulence-related properties that include initial adhesion and a high capacity for mucin degradation at the vaginal epithelial mucosal surface. Our results underline the importance of understanding the role of different anerobic bacteria in BV.
Importance: Bacterial vaginosis (BV) is a common vaginal infection that affects a significant proportion of women and is associated with reduced fertility and increased risk of secondary infections. Gardnerella vaginalis is the most well-known BV-associated bacterium, but Prevotella species including P. timonensis and P. bivia may also play an important role. We showed that, similar to G. vaginalis, P. timonensis adhered well to the vaginal epithelium, suggesting that both bacteria could be important in the first stage of infection. Compared to the other bacteria, P. timonensis was unique in efficiently removing the protective mucin sugars that cover the vaginal epithelium. These results underscore that vaginal bacteria play different roles in the initiation and development of BV.
Keywords: Hoylesella timonensis; NanH; Prevotella bivia; Prevotella timonensis; anerobic bacteria; fucosidase; mucus; neuraminidase; vaginal microbiota; α2,3-linked sialic acids; α2,6-linked sialic acids.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
-
- Cohen CR, Lingappa JR, Baeten JM, Ngayo MO, Spiegel CA, Hong T, Donnell D, Celum C, Kapiga S, Delany S, Bukusi EA. 2012. Bacterial vaginosis associated with increased risk of female-to-male HIV-1 transmission: a prospective cohort analysis among African couples. PLoS Med 9:e1001251. doi: 10.1371/journal.pmed.1001251 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
