

Orthologs of Plasmodium ICM1 are dispensable for Ca2+ mobilization in Toxoplasma gondii
- PMID: 39162502
- PMCID: PMC11448412
- DOI: 10.1128/spectrum.01229-24
Orthologs of Plasmodium ICM1 are dispensable for Ca2+ mobilization in Toxoplasma gondii
Abstract
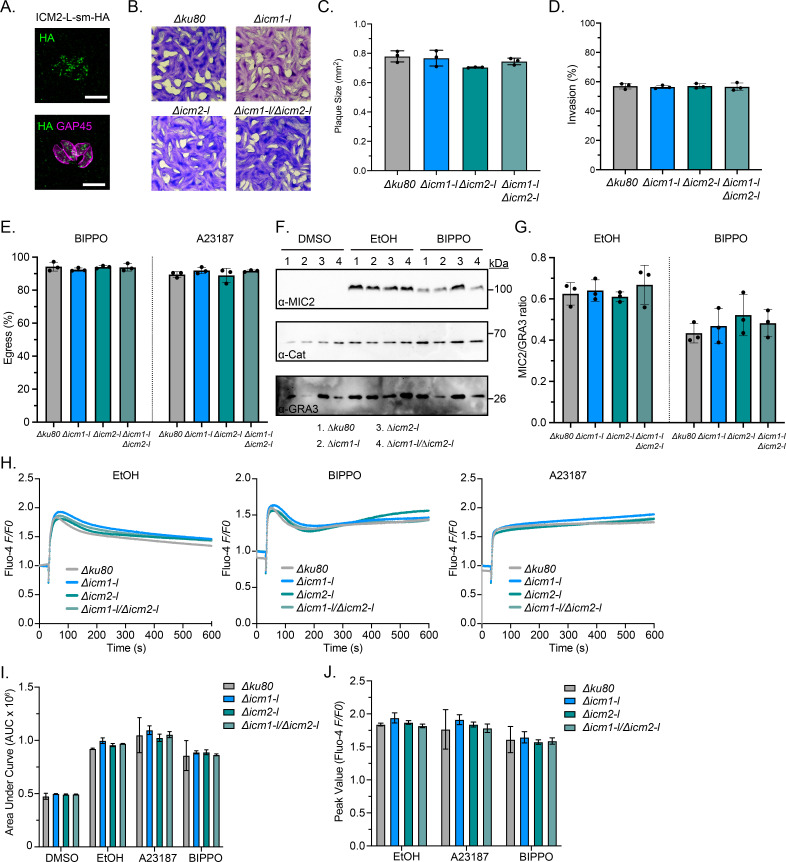
Apicomplexan parasites mobilize ionic calcium (Ca2+) from intracellular stores to promote microneme secretion and facilitate motile processes including gliding motility, invasion, and egress. Recently, a multipass transmembrane protein, ICM1, was found to be
Keywords: Apicomplexa; Plasmodium; Toxoplasma; Toxoplasma gondii; apicomplexan; cAMP; cGMP; calcium; calcium flux; calcium signaling; cyclic GMP; motility.
Conflict of interest statement
The authors declare no conflict of interest.
Figures



References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
