

H-NS is a bacterial transposon capture protein
- PMID: 39164300
- PMCID: PMC11335895
- DOI: 10.1038/s41467-024-51407-5
H-NS is a bacterial transposon capture protein
Abstract
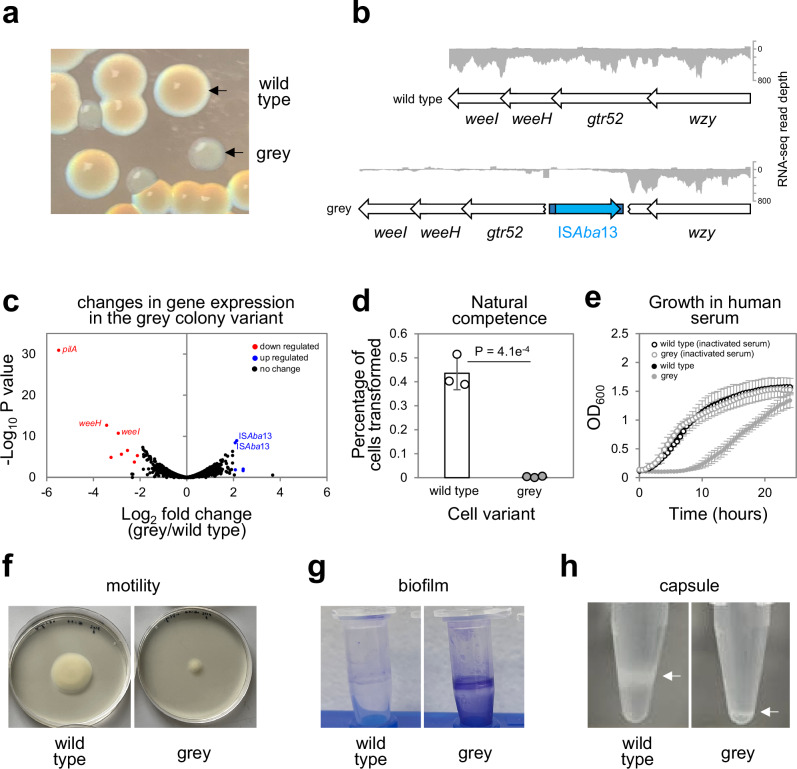
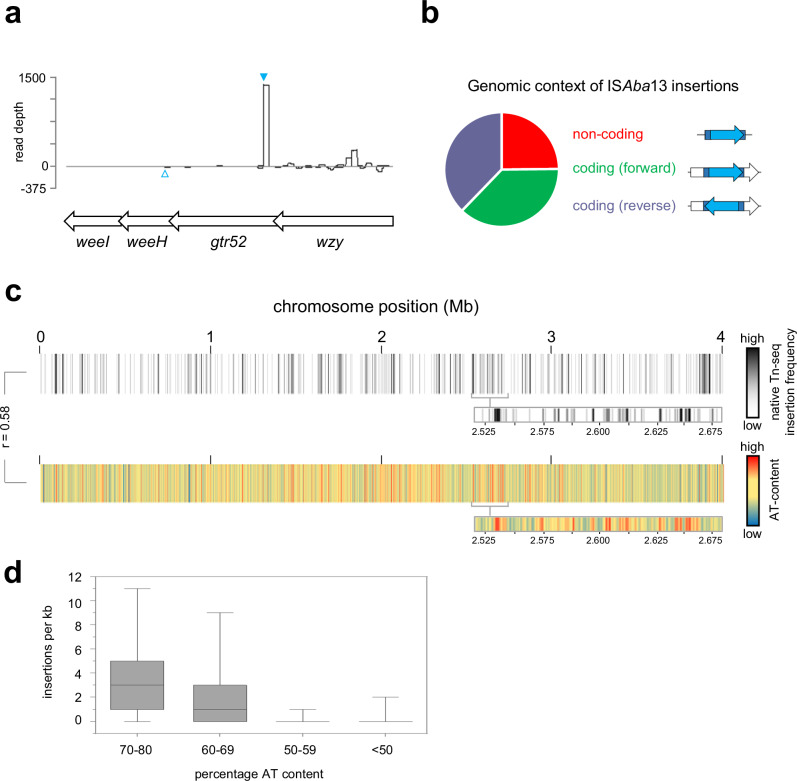
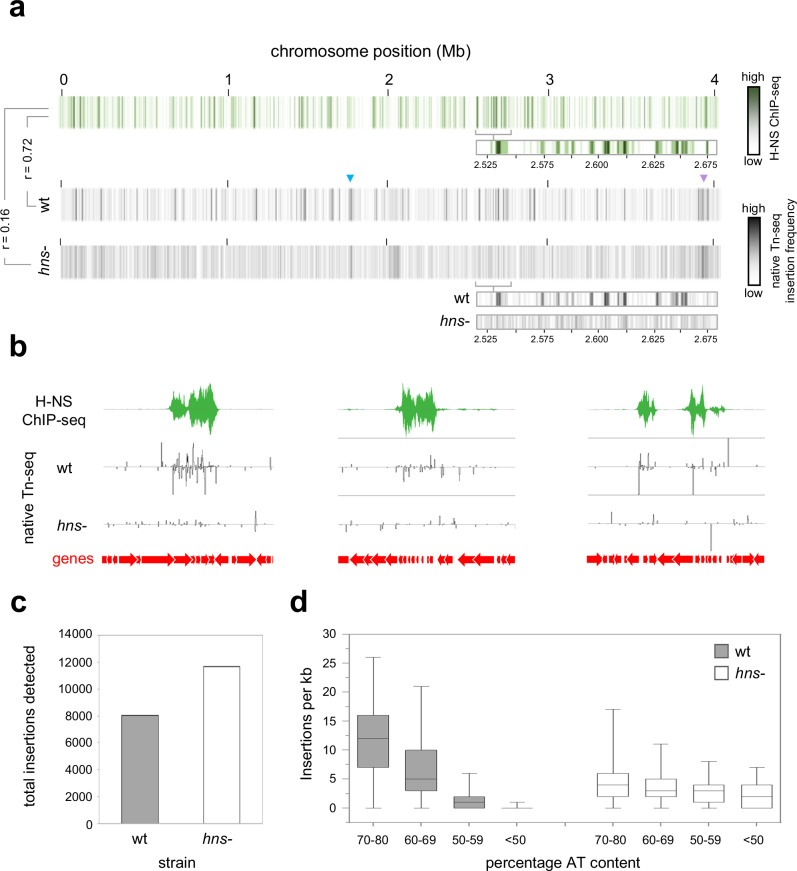
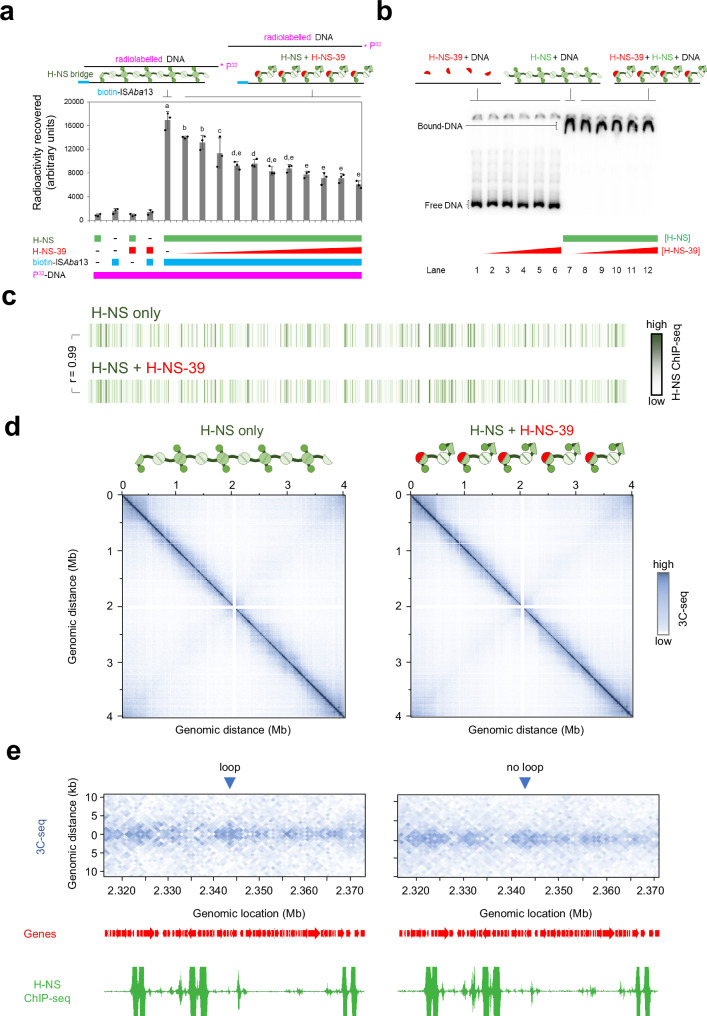
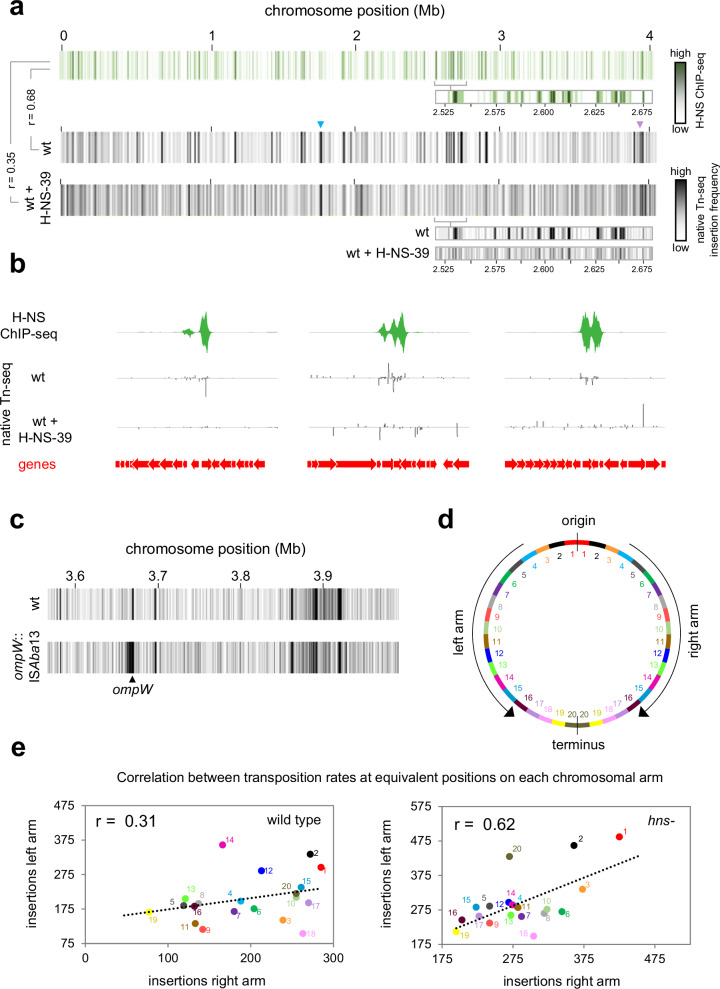
The histone-like nucleoid structuring (H-NS) protein is a DNA binding factor, found in gammaproteobacteria, with functional equivalents in diverse microbes. Universally, such proteins are understood to silence transcription of horizontally acquired genes. Here, we identify transposon capture as a major overlooked function of H-NS. Using genome-scale approaches, we show that H-NS bound regions are transposition "hotspots". Since H-NS often interacts with pathogenicity islands, such targeting creates clinically relevant phenotypic diversity. For example, in Acinetobacter baumannii, we identify altered motility, biofilm formation, and interactions with the human immune system. Transposon capture is mediated by the DNA bridging activity of H-NS and, if absent, more ubiquitous transposition results. Consequently, transcribed and essential genes are disrupted. Hence, H-NS directs transposition to favour evolutionary outcomes useful for the host cell.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
