

Smooth muscle-specific deletion of cellular communication network factor 2 causes severe aorta malformation and atherosclerosis
- PMID: 39167826
- PMCID: PMC11630017
- DOI: 10.1093/cvr/cvae174
Smooth muscle-specific deletion of cellular communication network factor 2 causes severe aorta malformation and atherosclerosis
Abstract
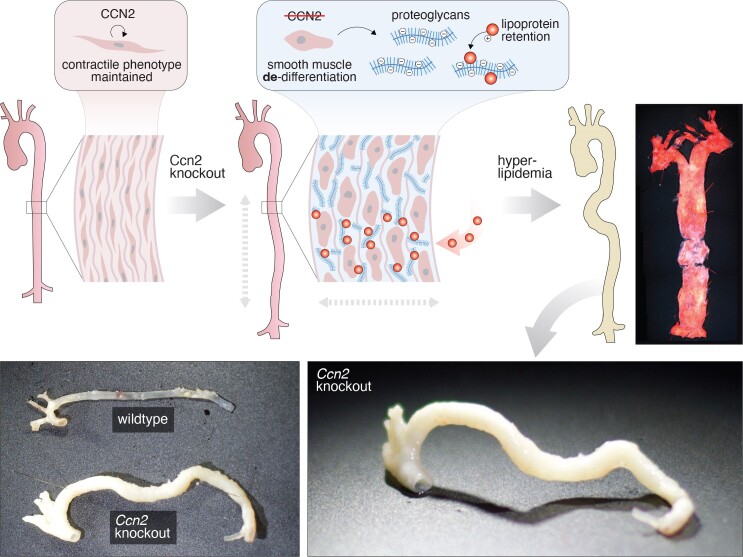
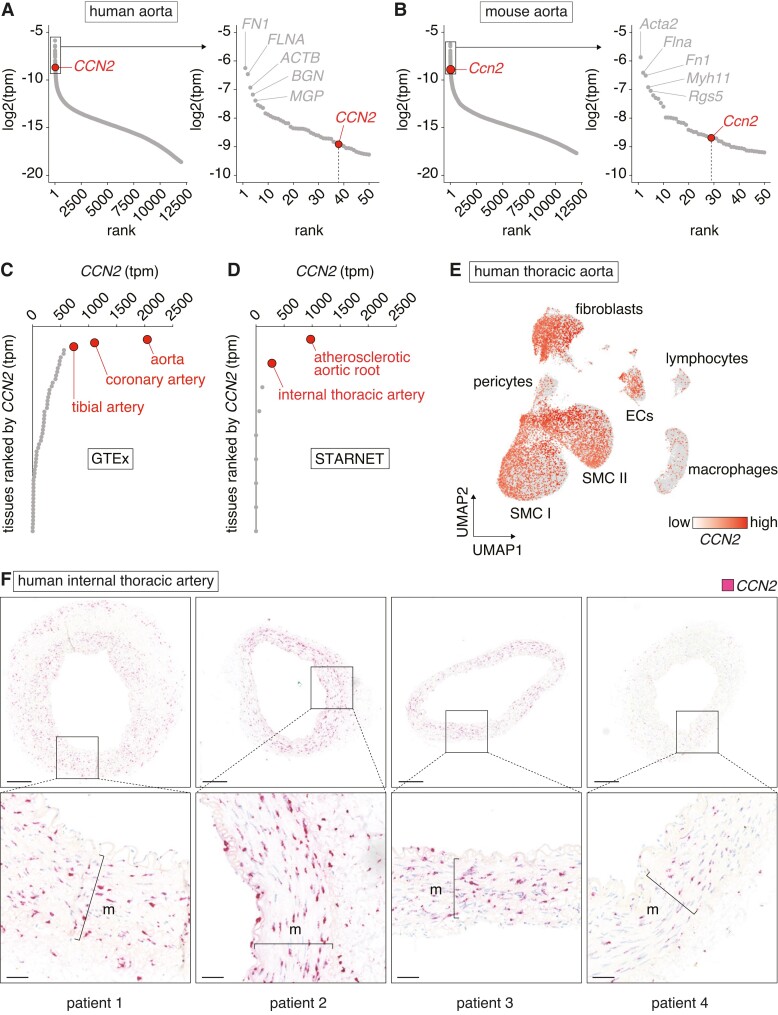
Aims: Cellular communication network factor 2 (CCN2) is a matricellular protein implicated in fibrotic diseases, with ongoing clinical trials evaluating anti-CCN2-based therapies. By uncovering CCN2 as abundantly expressed in non-diseased artery tissue, this study aimed to investigate the hypothesis that CCN2 plays a pivotal role in maintaining smooth muscle cell (SMC) phenotype and protection against atherosclerosis.
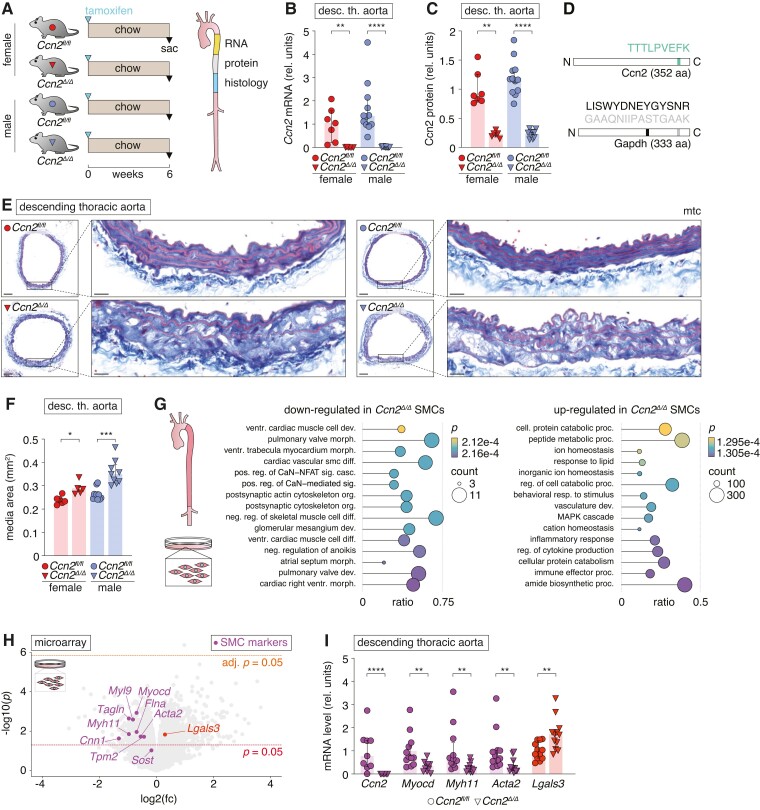
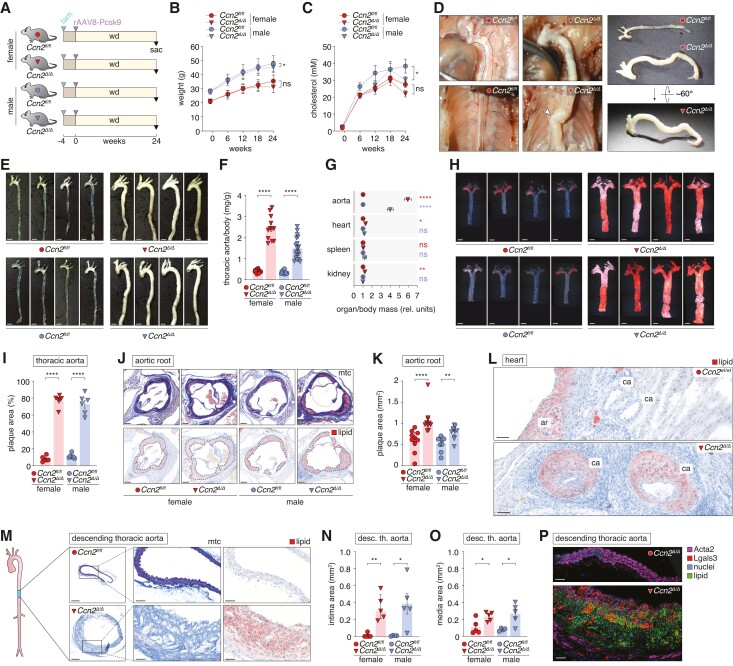
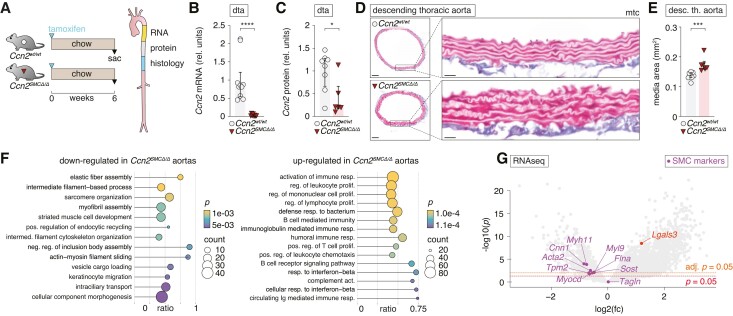
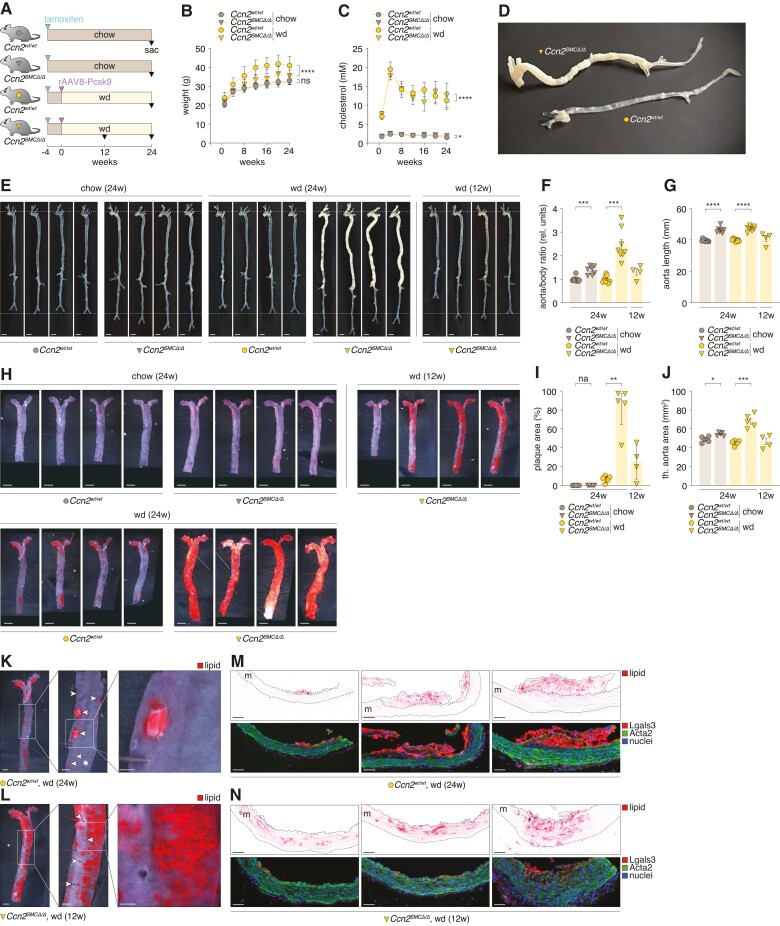
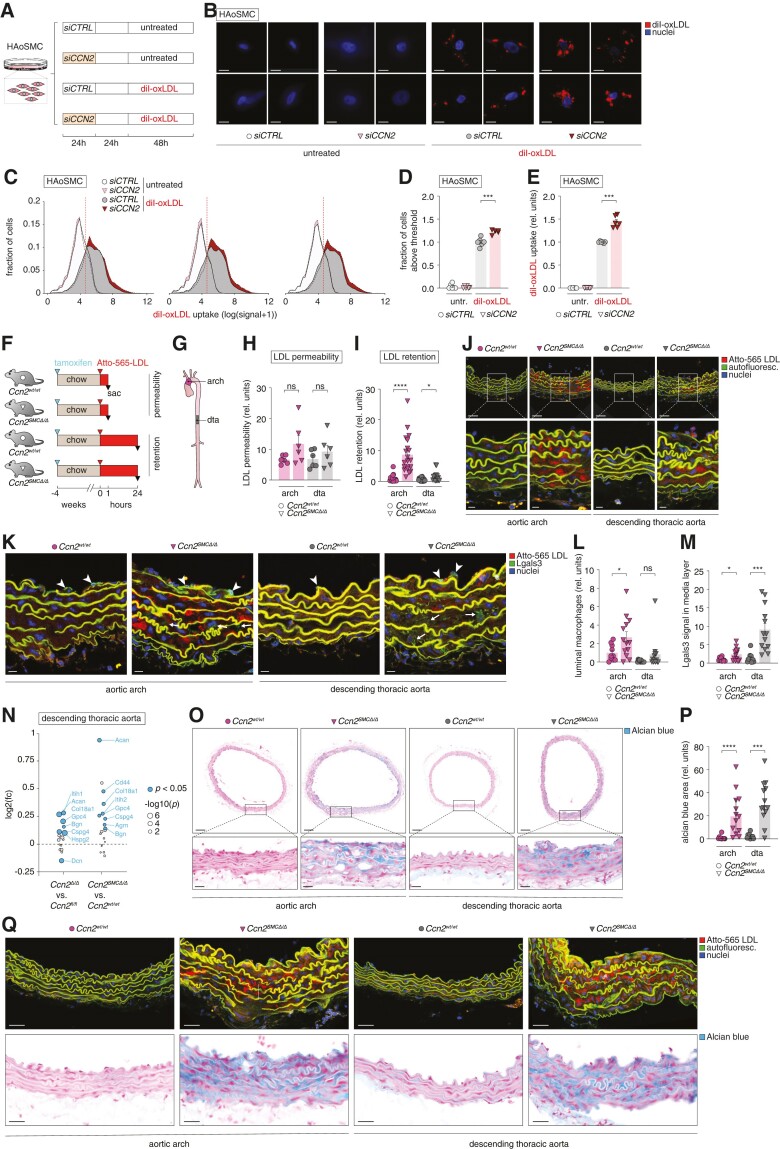
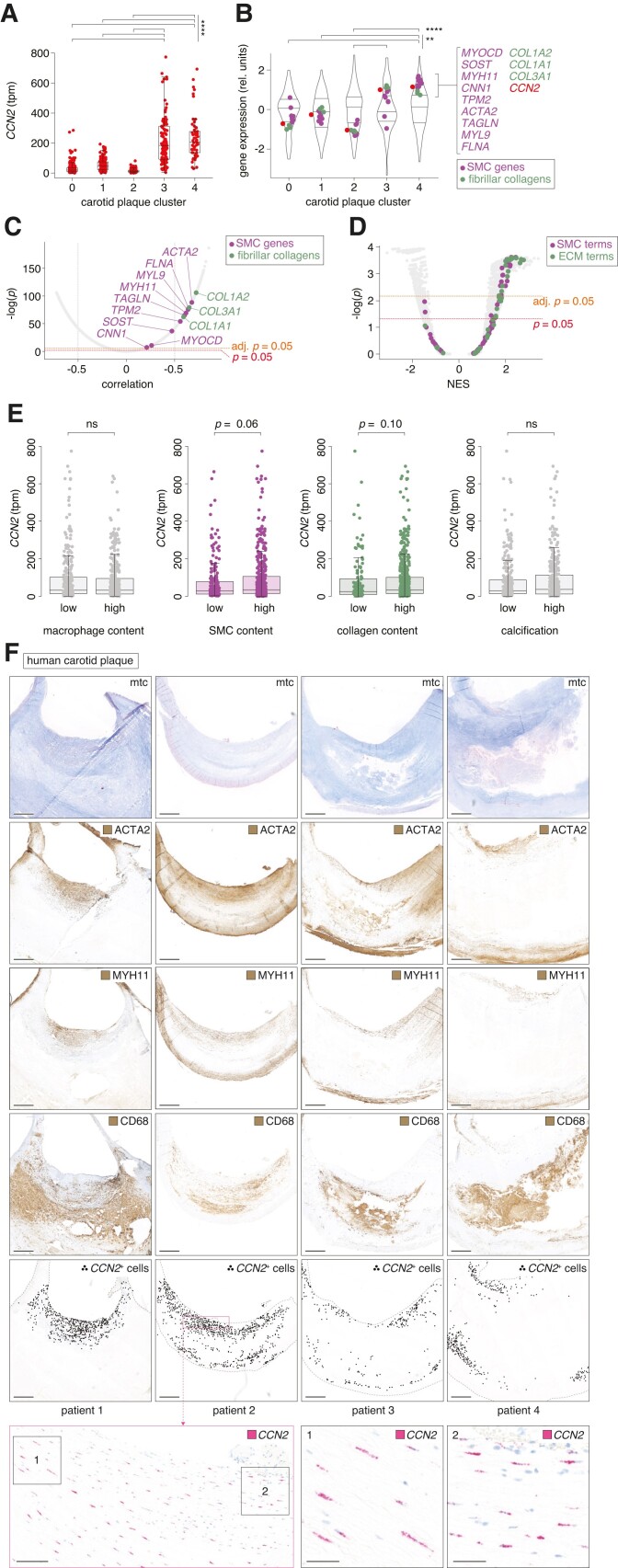
Methods and results: Global- and SMC-specific Ccn2 knockout mouse models were employed to demonstrate that Ccn2 deficiency leads to SMC de-differentiation, medial thickening, and aorta elongation under normolipidaemic conditions. Inducing hyperlipidaemia in both models resulted in severe aorta malformation and a 17-fold increase in atherosclerosis formation. Lipid-rich lesions developed at sites of the vasculature typically protected from atherosclerosis development by laminar blood flow, covering 90% of aortas and extending to other vessels, including coronary arteries. Evaluation at earlier time points revealed medial lipid accumulation as a lesion-initiating event. Fluorescently labelled LDL injection followed by confocal microscopy showed increased LDL retention in the medial layer of Ccn2 knockout aortas, likely attributed to marked proteoglycan enrichment of the medial extracellular matrix. Analyses leveraging data from the Athero-Express study cohort indicated the relevance of CCN2 in established human lesions, as CCN2 correlated with SMC marker transcripts across 654 transcriptomically profiled carotid plaques. These findings were substantiated through in situ hybridization showing CCN2 expression predominantly in the fibrous cap.
Conclusion: This study identifies CCN2 as a major constituent of the normal artery wall, critical in regulating SMC differentiation and aorta integrity and possessing a protective role against atherosclerosis development. These findings underscore the need for further investigation into the potential effects of anti-CCN2-based therapies on the vasculature.
Keywords: Aorta; Atherosclerosis; Cellular communication network factor 2 (CCN2); Smooth muscle cell.
© The Author(s) 2024. Published by Oxford University Press on behalf of the European Society of Cardiology.
Conflict of interest statement
Conflict of interest: none declared.
Figures
References
-
- Borén J, Williams KJ. The central role of arterial retention of cholesterol-rich apolipoprotein-B-containing lipoproteins in the pathogenesis of atherosclerosis: a triumph of simplicity. Curr Opin Lipidol 2016;27:473–483. - PubMed
-
- Libby P, Buring JE, Badimon L, Hansson GK, Deanfield J, Bittencourt MS, Tokgözoğlu L, Lewis EF. Atherosclerosis. Nat Rev Dis Primers 2019;5:56. - PubMed
-
- Bentzon JF, Otsuka F, Virmani R, Falk E. Mechanisms of plaque formation and rupture. Circ Res 2014;114:1852–1866. - PubMed
-
- Pipes GC, Creemers EE, Olson EN. The myocardin family of transcriptional coactivators: versatile regulators of cell growth, migration, and myogenesis. Genes Dev 2006;20:1545–1556. - PubMed
-
- Basatemur GL, Jørgensen HF, Clarke MCH, Bennett MR, Mallat Z. Vascular smooth muscle cells in atherosclerosis. Nat Rev Cardiol 2019;16:727–744. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
