

A class I PI3K signalling network regulates primary cilia disassembly in normal physiology and disease
- PMID: 39168978
- PMCID: PMC11339396
- DOI: 10.1038/s41467-024-51354-1
A class I PI3K signalling network regulates primary cilia disassembly in normal physiology and disease
Abstract
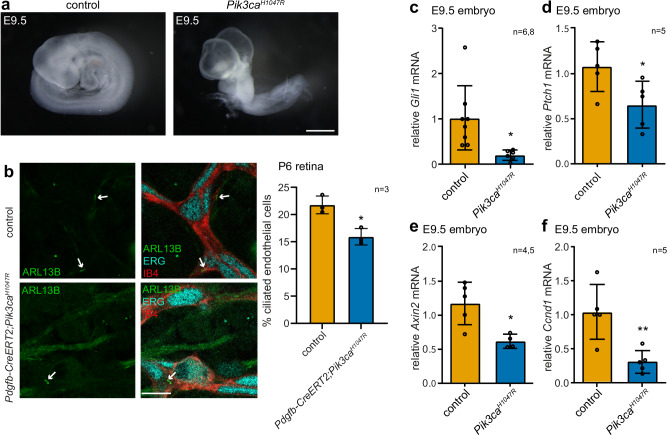
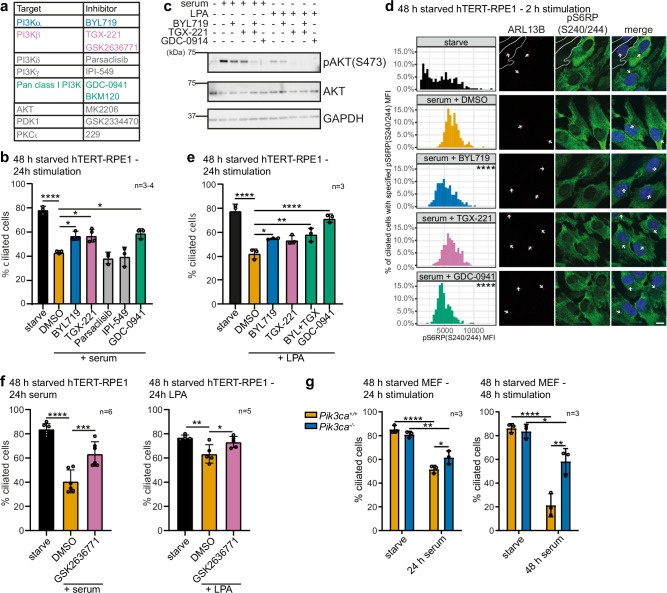
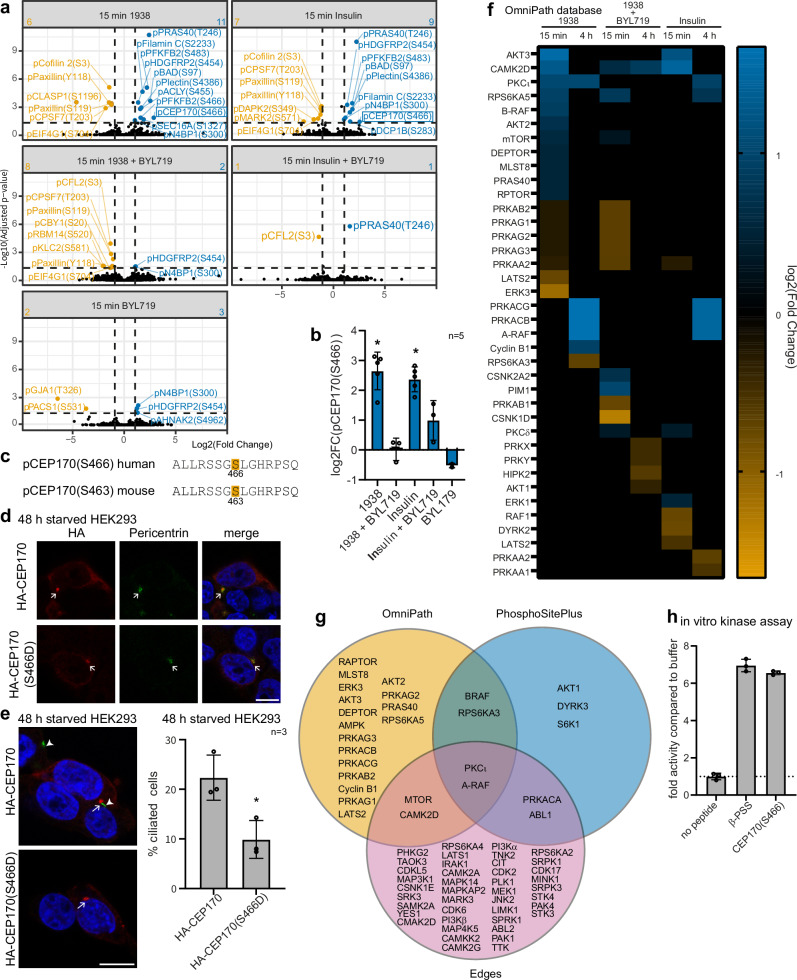
Primary cilia are antenna-like organelles which sense extracellular cues and act as signalling hubs. Cilia dysfunction causes a heterogeneous group of disorders known as ciliopathy syndromes affecting most organs. Cilia disassembly, the process by which cells lose their cilium, is poorly understood but frequently observed in disease and upon cell transformation. Here, we uncover a role for the PI3Kα signalling enzyme in cilia disassembly. Genetic PI3Kα-hyperactivation, as observed in PIK3CA-related overgrowth spectrum (PROS) and cancer, induced a ciliopathy-like phenotype during mouse development. Mechanistically, PI3Kα and PI3Kβ produce the PIP3 lipid at the cilia transition zone upon disassembly stimulation. PI3Kα activation initiates cilia disassembly through a kinase signalling axis via the PDK1/PKCι kinases, the CEP170 centrosomal protein and the KIF2A microtubule-depolymerising kinesin. Our data suggest diseases caused by PI3Kα-activation may be considered 'Disorders with Ciliary Contributions', a recently-defined subset of ciliopathies in which some, but not all, of the clinical manifestations result from cilia dysfunction.
© 2024. The Author(s).
Conflict of interest statement
B.V. is a consultant for iOnctura (Geneva, Switzerland), Pharming (Leiden, the Netherlands) and shareholder of Open Orphan and Poolbeg Pharma (Dublin, Ireland), P.J.P. is a consultant for Apollo Therapeutics and Phoremost. P.R.C. is a co-founder and director of Kinomica Ltd, a company active in the area of phosphoproteomic diagnostics. The remaining authors declare no competing interests.
Figures




References
-
- Lovera, M. & Lüders, J. The ciliary impact of nonciliary gene mutations. Trends Cell Biol.31, 876–887 (2021). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
