

Palmitoylation of ULK1 by ZDHHC13 plays a crucial role in autophagy
- PMID: 39169022
- PMCID: PMC11339336
- DOI: 10.1038/s41467-024-51402-w
Palmitoylation of ULK1 by ZDHHC13 plays a crucial role in autophagy
Erratum in
-
Author Correction: Palmitoylation of ULK1 by ZDHHC13 plays a crucial role in autophagy.Nat Commun. 2024 Oct 3;15(1):8556. doi: 10.1038/s41467-024-52646-2. Nat Commun. 2024. PMID: 39362856 Free PMC article. No abstract available.
Abstract
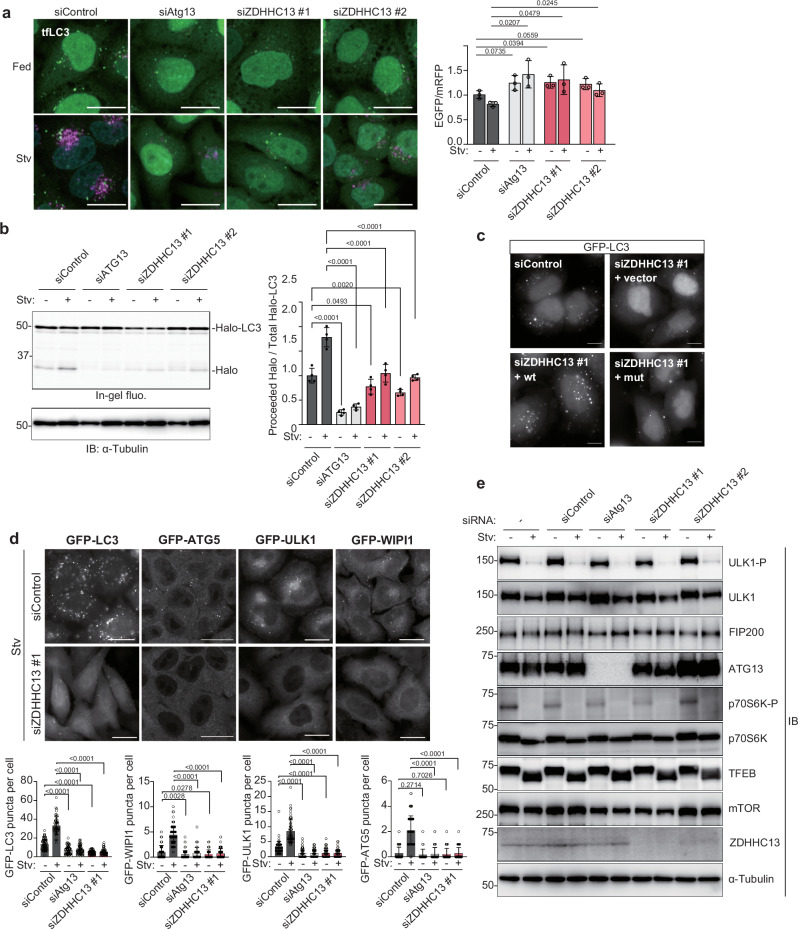
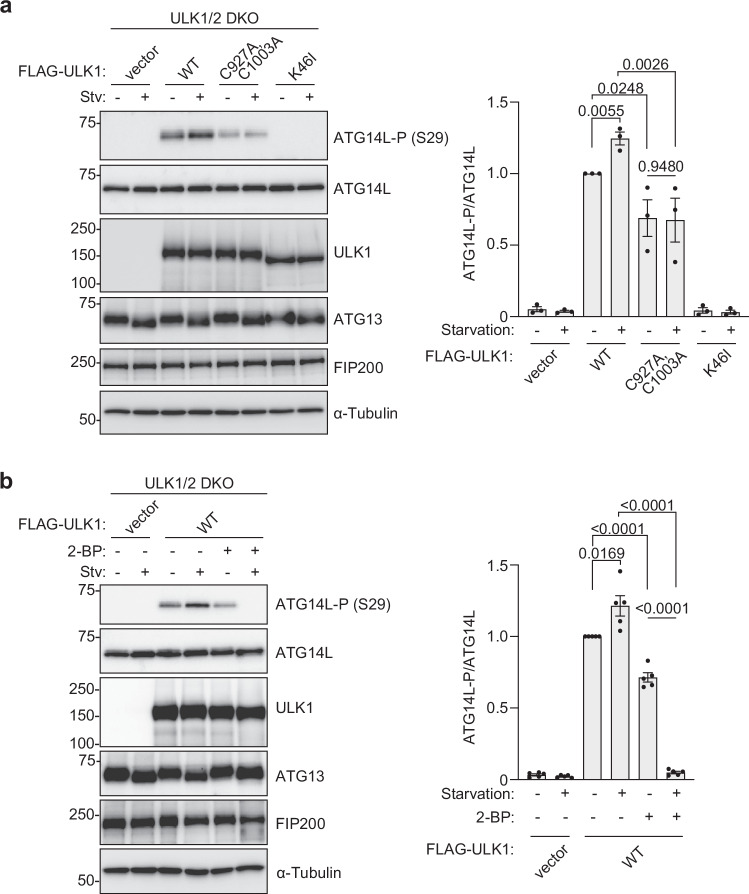
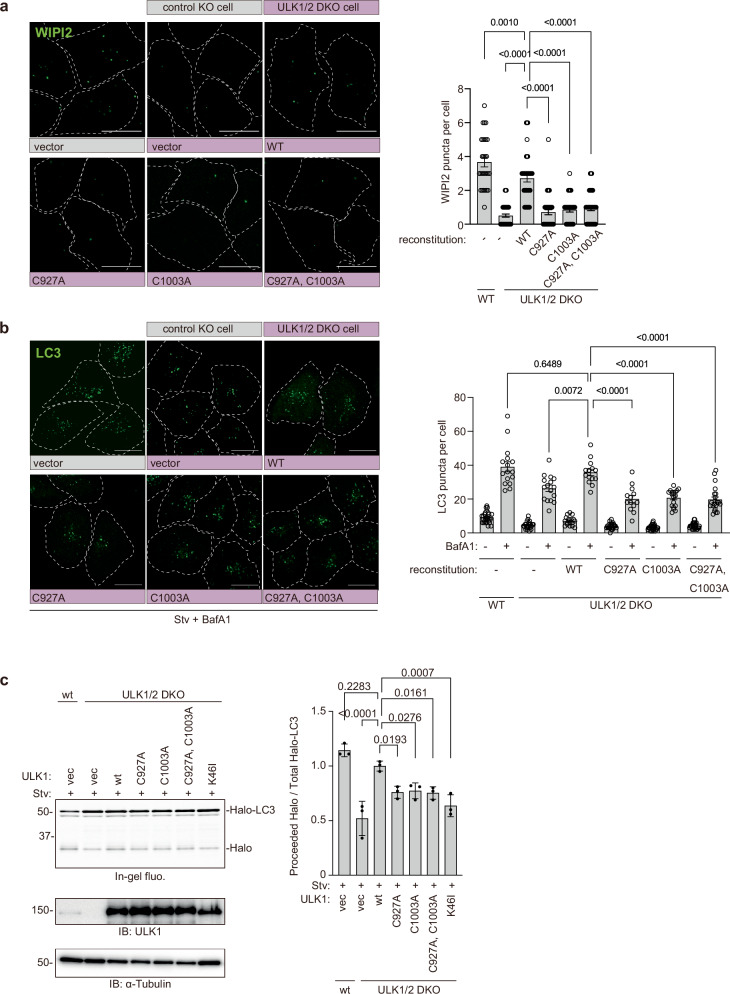
Autophagy is a highly conserved process from yeast to mammals in which intracellular materials are engulfed by a double-membrane organelle called autophagosome and degrading materials by fusing with the lysosome. The process of autophagy is regulated by sequential recruitment and function of autophagy-related (Atg) proteins. Genetic hierarchical analyses show that the ULK1 complex comprised of ULK1-FIP200-ATG13-ATG101 translocating from the cytosol to autophagosome formation sites as a most upstream ATG factor; this translocation is critical in autophagy initiation. However, how this translocation occurs remains unclear. Here, we show that ULK1 is palmitoylated by palmitoyltransferase ZDHHC13 and translocated to the autophagosome formation site upon autophagy induction. We find that the ULK1 palmitoylation is required for autophagy initiation. Moreover, the ULK1 palmitoylated enhances the phosphorylation of ATG14L, which is required for activating PI3-Kinase and producing phosphatidylinositol 3-phosphate, one of the autophagosome membrane's lipids. Our results reveal how the most upstream ULK1 complex translocates to the autophagosome formation sites during autophagy.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
MeSH terms
Substances
Grants and funding
- JP15H04371/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP21K06152/MEXT | Japan Society for the Promotion of Science (JSPS)
- 21K06169/MEXT | Japan Society for the Promotion of Science (JSPS)
- 20K05839/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP26251020/MEXT | Japan Society for the Promotion of Science (JSPS)
LinkOut - more resources
Full Text Sources
