Thermal infrared directs host-seeking behaviour in Aedes aegypti mosquitoes
- PMID: 39169183
- PMCID: PMC11410652
- DOI: 10.1038/s41586-024-07848-5
Thermal infrared directs host-seeking behaviour in Aedes aegypti mosquitoes
Abstract
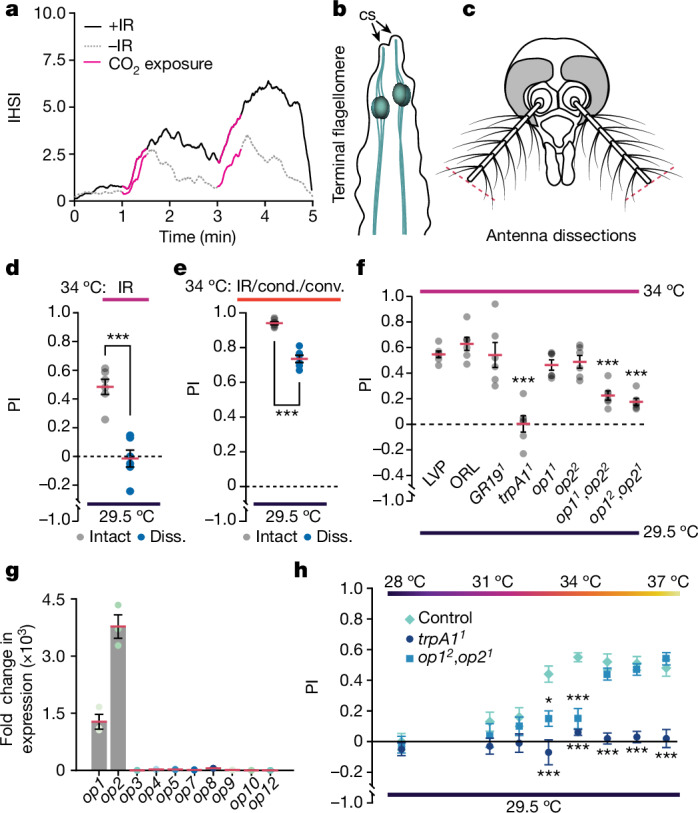
Mosquito-borne diseases affect hundreds of millions of people annually and disproportionately impact the developing world1,2. One mosquito species, Aedes aegypti, is a primary vector of viruses that cause dengue, yellow fever and Zika. The attraction of Ae. aegypti female mosquitos to humans requires integrating multiple cues, including CO2 from breath, organic odours from skin and visual cues, all sensed at mid and long ranges, and other cues sensed at very close range3-6. Here we identify a cue that Ae. aegypti use as part of their sensory arsenal to find humans. We demonstrate that Ae. aegypti sense the infrared (IR) radiation emanating from their targets and use this information in combination with other cues for highly effective mid-range navigation. Detection of thermal IR requires the heat-activated channel TRPA1, which is expressed in neurons at the tip of the antenna. Two opsins are co-expressed with TRPA1 in these neurons and promote the detection of lower IR intensities. We propose that radiant energy causes local heating at the end of the antenna, thereby activating temperature-sensitive receptors in thermosensory neurons. The realization that thermal IR radiation is an outstanding mid-range directional cue expands our understanding as to how mosquitoes are exquisitely effective in locating hosts.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures