

FYN-mediated phosphorylation of BCKDK at Y151 promotes GBM proliferation by increasing the oncogenic metabolite N-acetyl-L-alanine
- PMID: 39170503
- PMCID: PMC11336342
- DOI: 10.1016/j.heliyon.2024.e33663
FYN-mediated phosphorylation of BCKDK at Y151 promotes GBM proliferation by increasing the oncogenic metabolite N-acetyl-L-alanine
Abstract
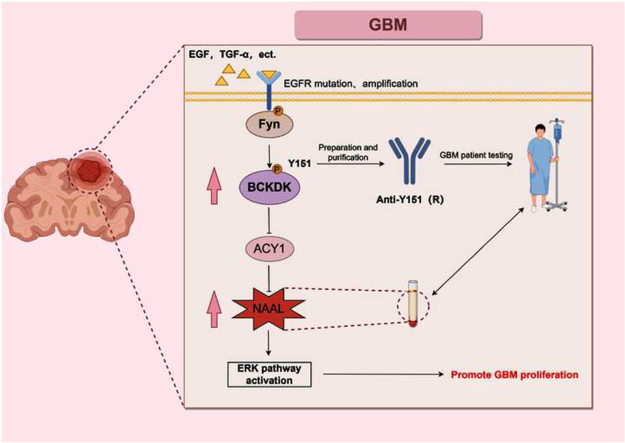
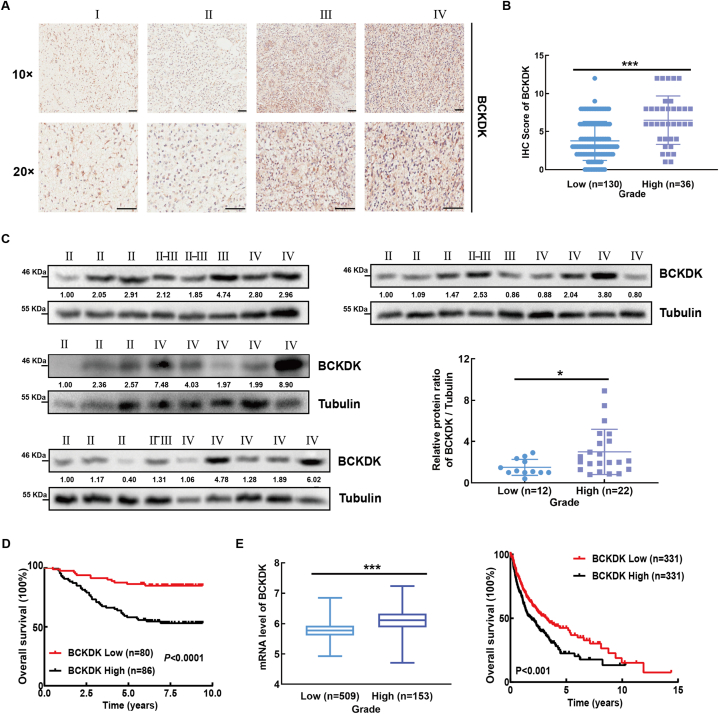
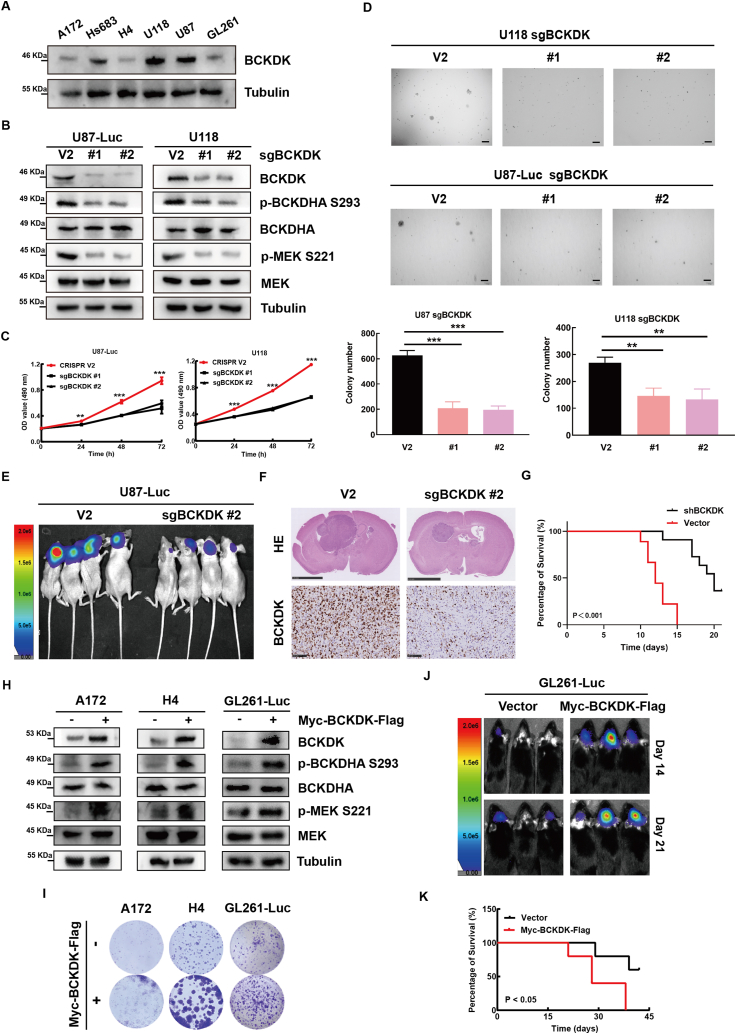
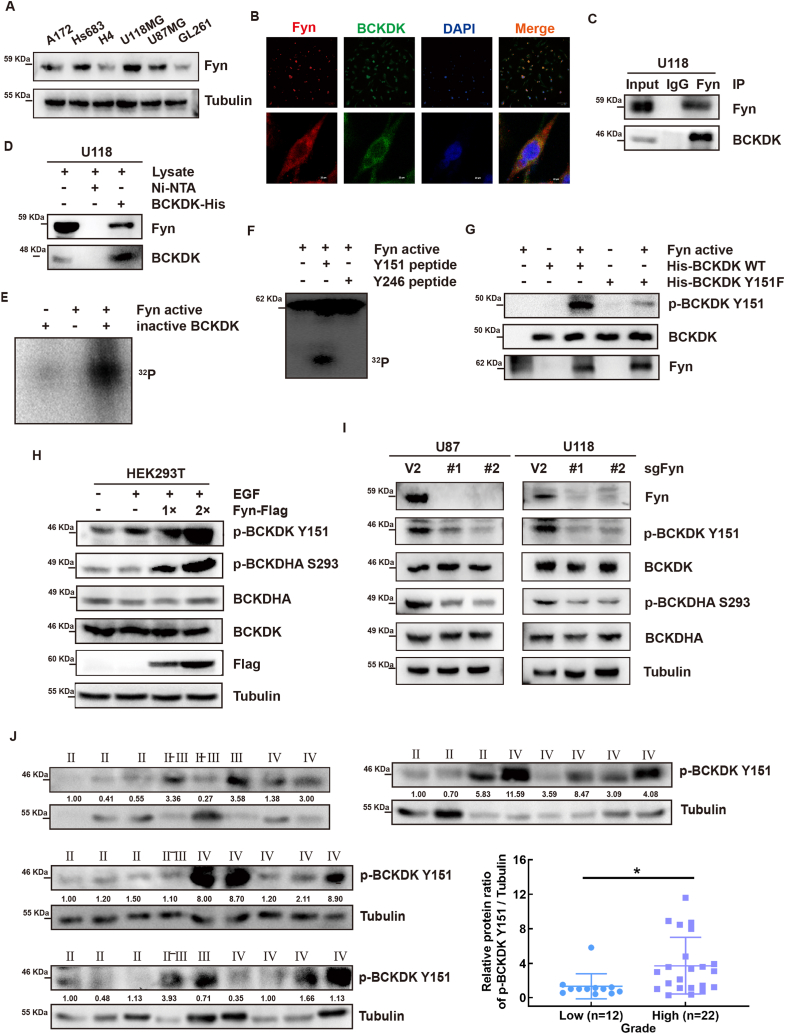
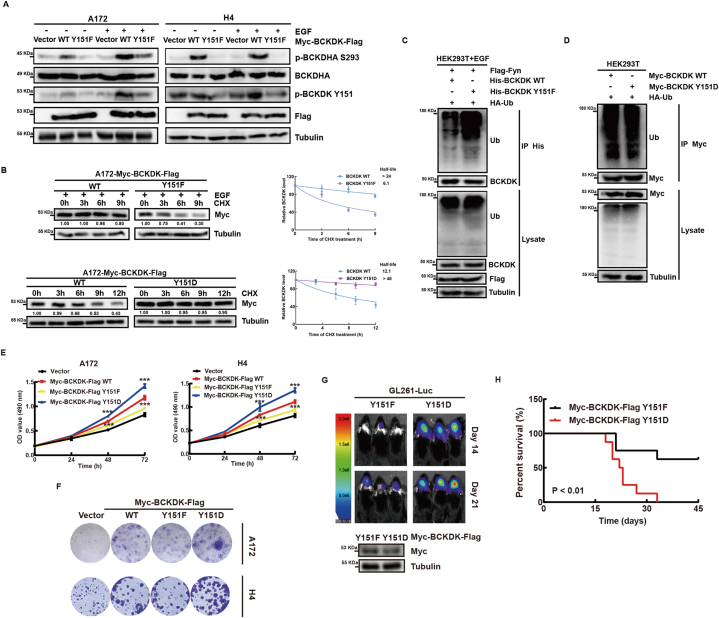
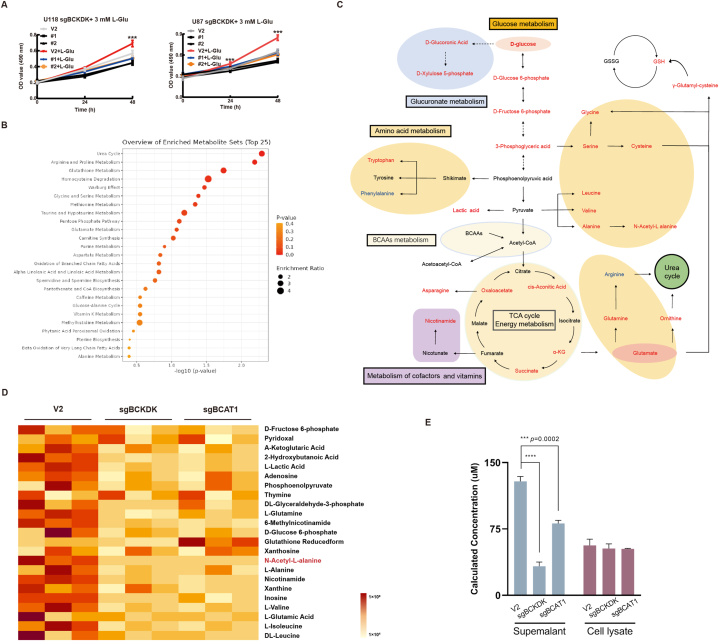
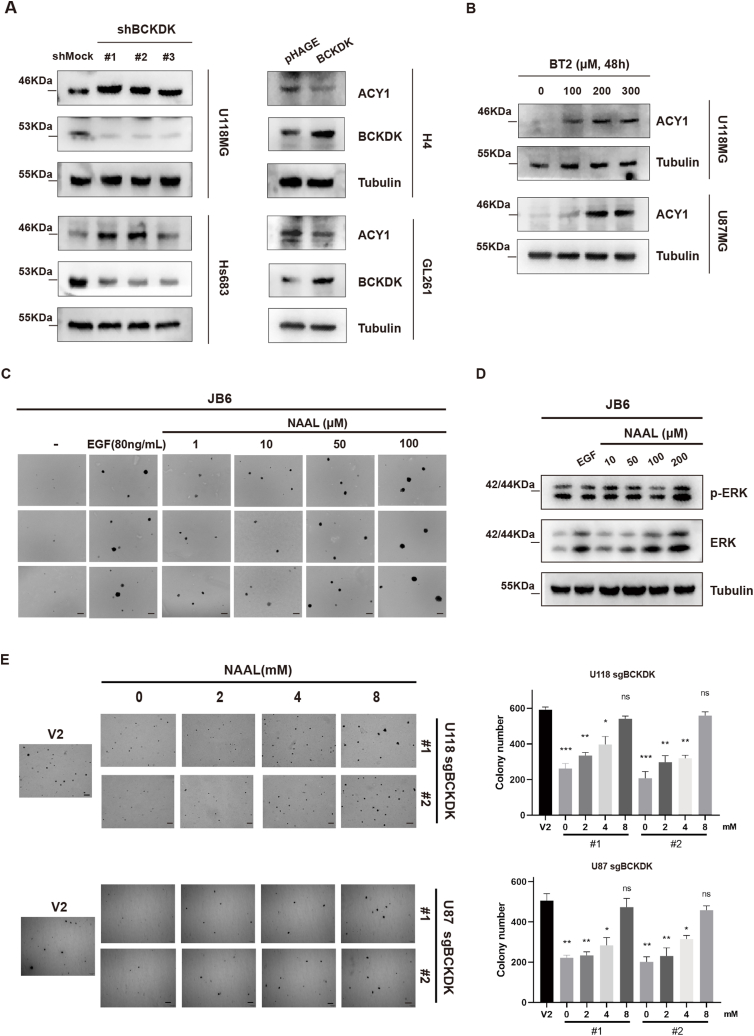
Branched chain α-keto acid dehydrogenase kinase (BCKDK) is a key enzyme involved in the metabolism of branched-chain amino acids (BCAAs). Its potential as a therapeutic target and prognostic factor for a variety of cancers has been widely reported. In this study, we investigated the expression of BCKDK in clinical glioma samples and found that BCKDK was significantly overexpressed in glioblastoma (GBM) and was associated with its poor prognosis. We further found that BCKDK is phosphorylated by tyrosine protein kinase Fyn at Y151, which increases its catalytic activity and stability, and demonstrate through in vivo and in vitro experiments that BCKDK phosphorylation promotes GBM cell proliferation. In addition, we found that the levels of the metabolite N-acetyl-L-alanine (NAAL) in GBM cells with high BCKDK were higher than those in the silencing group, and silencing or inhibition of BCKDK promotes the expression of ACY1, an enzyme that catalyzes the hydrolysis of NAAL into acetic acid and alanine. Exogenous addition of NAAL can activate the ERK signaling pathway and promote the proliferation of GBM cells. Taken together, we identified a novel mechanism of BCKDK activation and found NAAL is a novel oncogenic metabolite. Our study confirms the importance of the Fyn-BCKDK-ACY1-NAAL signalling axis in the development of GBM and suggests that p-BCKDK (Y151) and NAAL can serve as potential predictors of GBM progression and prognosis.
Keywords: ACY1; BCKDK; Fyn; Glioblastoma; N-acetyl-L-alanine.
© 2024 The Authors.
Conflict of interest statement
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests:Feng Zhu reports financial support was provided by National Natural Science Foundation of China. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures
Similar articles
-
Cross-talk between BCKDK-mediated phosphorylation and STUB1-dependent ubiquitination degradation of BCAT1 promotes GBM progression.Cancer Lett. 2024 Jun 1;591:216849. doi: 10.1016/j.canlet.2024.216849. Epub 2024 Apr 16. Cancer Lett. 2024. PMID: 38621458
-
Phosphorylation of BCKDK of BCAA catabolism at Y246 by Src promotes metastasis of colorectal cancer.Oncogene. 2020 May;39(20):3980-3996. doi: 10.1038/s41388-020-1262-z. Epub 2020 Apr 1. Oncogene. 2020. PMID: 32238881 Free PMC article.
-
BCKDK of BCAA Catabolism Cross-talking With the MAPK Pathway Promotes Tumorigenesis of Colorectal Cancer.EBioMedicine. 2017 Jun;20:50-60. doi: 10.1016/j.ebiom.2017.05.001. Epub 2017 May 4. EBioMedicine. 2017. PMID: 28501528 Free PMC article.
-
The Role of Branched-Chain Amino Acids and Branched-Chain α-Keto Acid Dehydrogenase Kinase in Metabolic Disorders.Front Nutr. 2022 Jul 18;9:932670. doi: 10.3389/fnut.2022.932670. eCollection 2022. Front Nutr. 2022. PMID: 35923208 Free PMC article. Review.
-
The role of branched chain amino acids metabolic disorders in tumorigenesis and progression.Biomed Pharmacother. 2022 Sep;153:113390. doi: 10.1016/j.biopha.2022.113390. Epub 2022 Jul 14. Biomed Pharmacother. 2022. PMID: 36076478 Review.
Cited by
-
Unveiling the Maze: Branched-Chain Amino Acids Fueling the Dynamics of Cancer Metabolism and Progression.Cancers (Basel). 2025 May 23;17(11):1751. doi: 10.3390/cancers17111751. Cancers (Basel). 2025. PMID: 40507232 Free PMC article. Review.
References
-
- Ohgaki H., Kleihues P. Population-based studies on incidence, survival rates, and genetic alterations in astrocytic and oligodendroglial gliomas. J. Neuropathol. Exp. Neurol. 2005;64(6):479–489. - PubMed
-
- Louis D.N., et al. The 2016 World Health Organization classification of tumors of the central nervous system: a summary. Acta Neuropathol. 2016;131(6):803–820. - PubMed
-
- Wen P.Y., Reardon D.A. Neuro-oncology in 2015: progress in glioma diagnosis, classification and treatment. Nat. Rev. Neurol. 2016;12(2):69–70. - PubMed
-
- Bush N.A., Chang S.M., Berger M.S. Current and future strategies for treatment of glioma. Neurosurg. Rev. 2017;40(1):1–14. - PubMed
LinkOut - more resources
Full Text Sources
Miscellaneous
