

The nuclear export protein XPO1 provides a peptide ligand for natural killer cells
- PMID: 39178254
- PMCID: PMC11343027
- DOI: 10.1126/sciadv.ado6566
The nuclear export protein XPO1 provides a peptide ligand for natural killer cells
Abstract
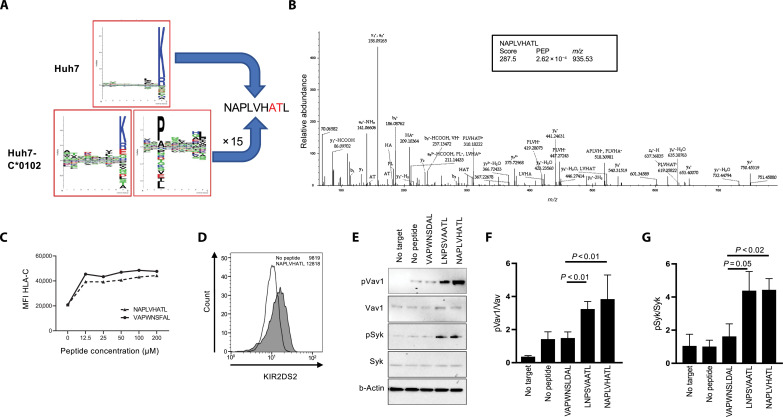
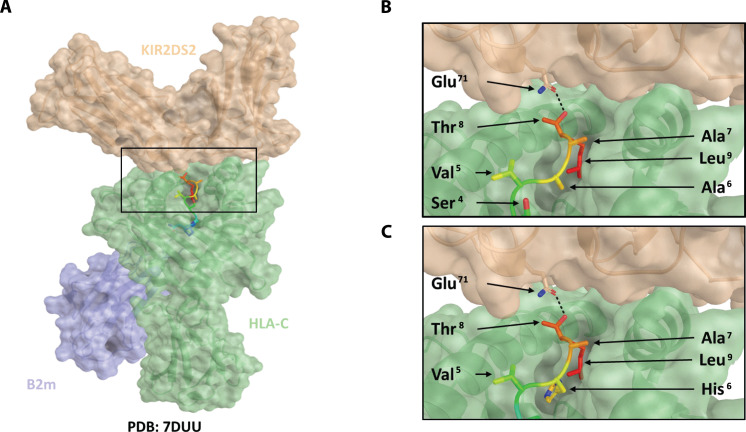
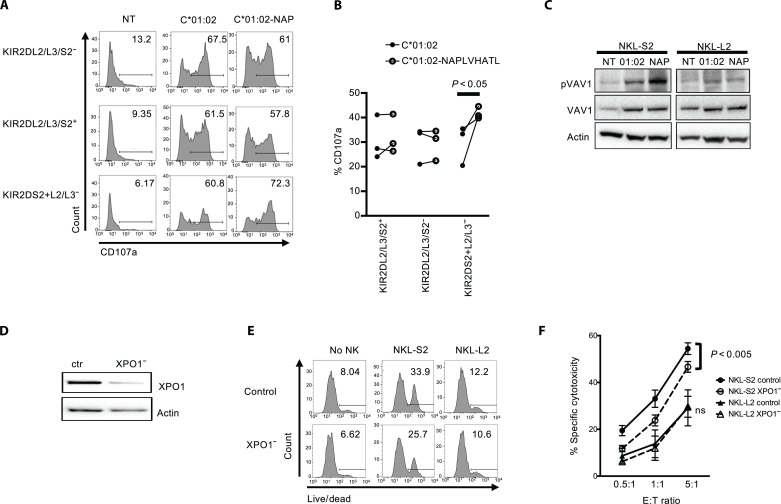
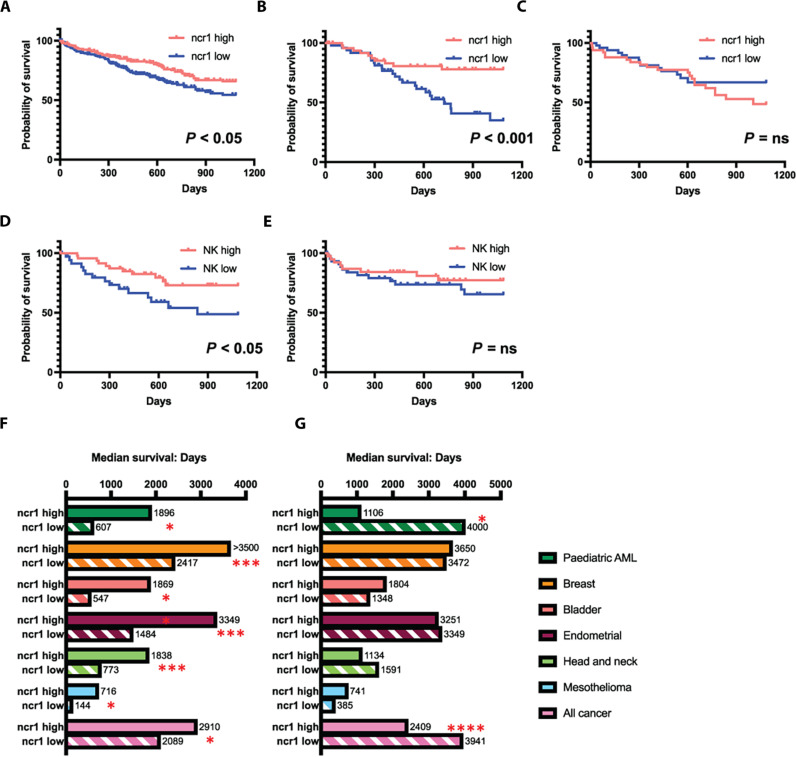
XPO1 (Exportin-1/CRM1) is a nuclear export protein that is frequently overexpressed in cancer and functions as a driver of oncogenesis. Currently small molecules that target XPO1 are being used in the clinic as anticancer agents. We identify XPO1 as a target for natural killer (NK) cells. Using immunopeptidomics, we have identified a peptide derived from XPO1 that can be recognized by the activating NK cell receptor KIR2DS2 in the context of human leukocyte antigen-C. The peptide can be endogenously processed and presented to activate NK cells specifically through this receptor. Although high XPO1 expression in cancer is commonly associated with a poor prognosis, we show that the outcome of specific cancers, such as hepatocellular carcinoma, can be substantially improved if there is concomitant evidence of NK cell infiltration. We thus identify XPO1 as a bona fide tumor antigen recognized by NK cells that offers an opportunity for a personalized approach to NK cell therapy for solid tumors.
Figures
References
-
- Chiossone L., Dumas P. Y., Vienne M., Vivier E., Natural killer cells and other innate lymphoid cells in cancer. Nat. Rev. Immunol. 18, 671–688 (2018). - PubMed
-
- Vilches C., Parham P., KIR: Diverse, rapidly evolving receptors of innate and adaptive immunity. Annu. Rev. Immunol. 20, 217–251 (2002). - PubMed
-
- Leone P., De Re V., Vacca A., Dammacco F., Racanelli V., Cancer treatment and the KIR-HLA system: An overview. Clin. Exp. Med. 17, 419–429 (2017). - PubMed
-
- Bao X., Hanson A. L., Madeleine M. M., Wang S. S., Schwartz S. M., Newell F., Pettersson-Kymmer U., Hemminki K., Tiews S., Steinberg W., Rader J. S., Castro F., Safaeian M., Franco E. L., Coutlee F., Ohlsson C., Cortes A., Marshall M., Mukhopadhyay P., Cremin K., Johnson L. G., Garland S. M., Tabrizi S. N., Wentzensen N., Sitas F., Trimble C., Little J., Cruickshank M., Frazer I. H., Hildesheim A., Brown M. A., Duncan E. L., Sun Y. P., Leo P. J., HLA and KIR associations of cervical neoplasia. J. Infect. Dis. 218, 2006–2015 (2018). - PMC - PubMed
-
- Cariani E., Pilli M., Zerbini A., Rota C., Olivani A., Zanelli P., Zanetti A., Trenti T., Ferrari C., Missale G., HLA and killer immunoglobulin-like receptor genes as outcome predictors of hepatitis C virus-related hepatocellular carcinoma. Clin. Cancer Res. 19, 5465–5473 (2013). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
