

Thyroid hormone remodels cortex to coordinate body-wide metabolism and exploration
- PMID: 39178853
- PMCID: PMC11455614
- DOI: 10.1016/j.cell.2024.07.041
Thyroid hormone remodels cortex to coordinate body-wide metabolism and exploration
Abstract
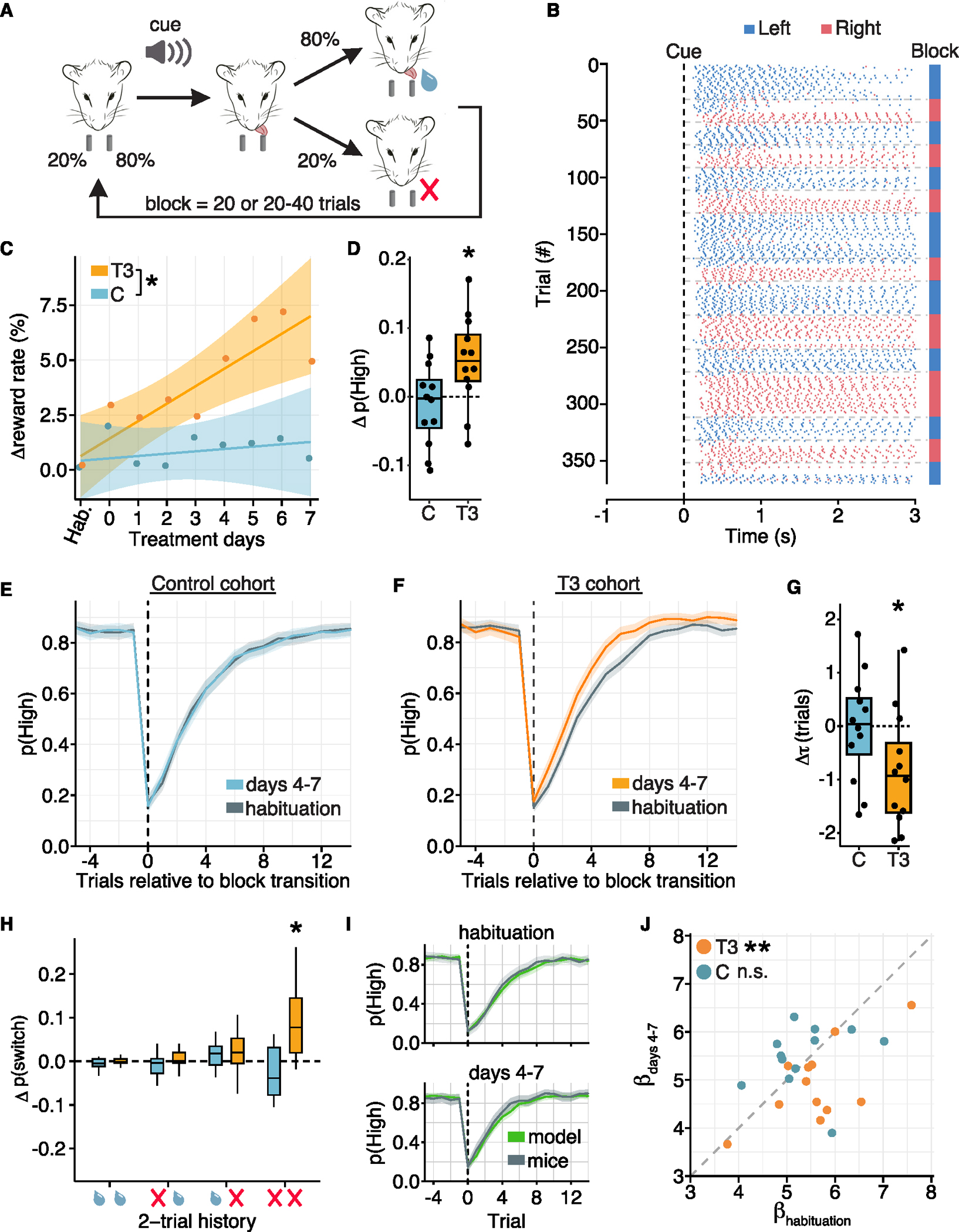
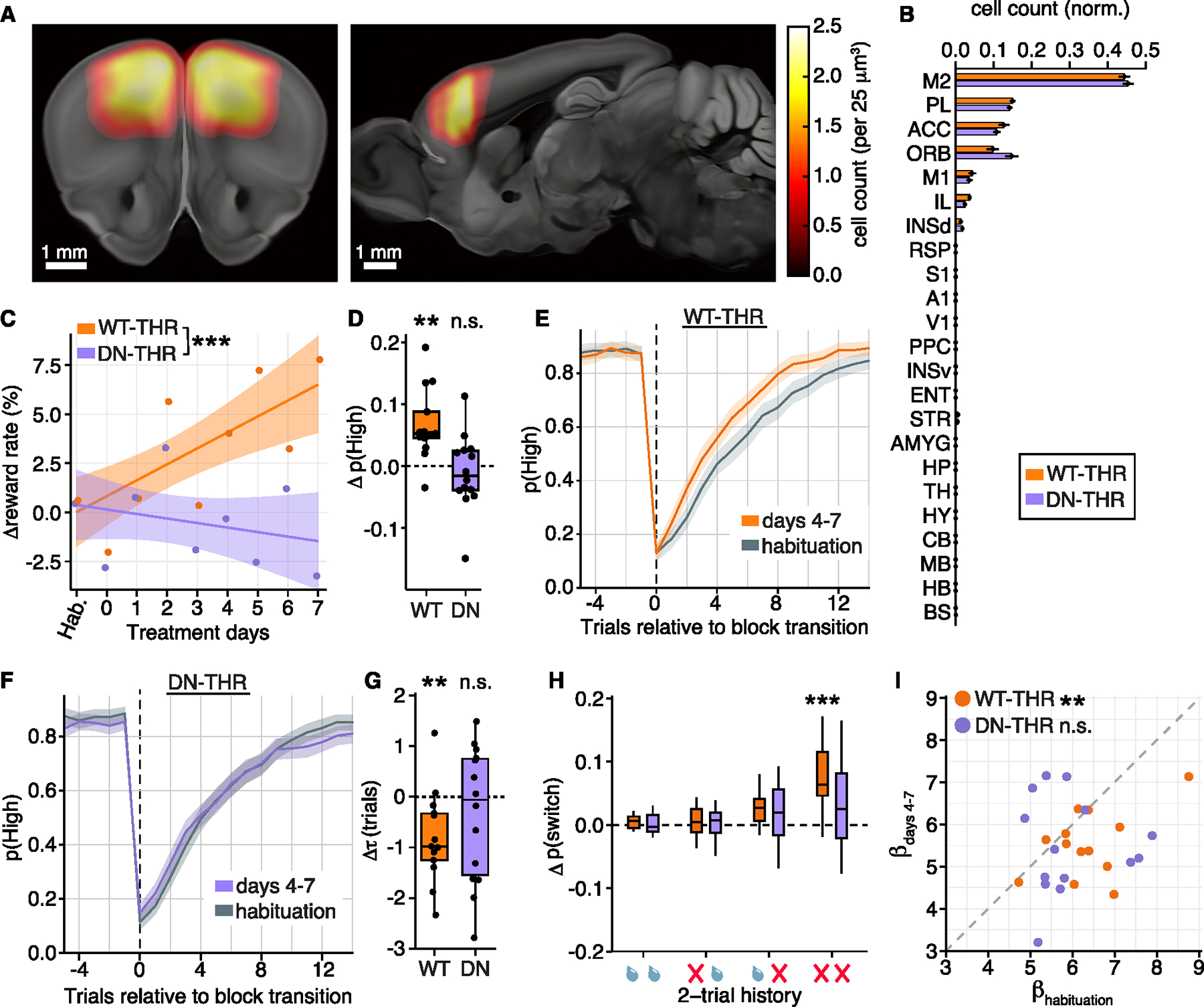
Animals adapt to environmental conditions by modifying the function of their internal organs, including the brain. To be adaptive, alterations in behavior must be coordinated with the functional state of organs throughout the body. Here, we find that thyroid hormone-a regulator of metabolism in many peripheral organs-directly activates cell-type-specific transcriptional programs in the frontal cortex of adult male mice. These programs are enriched for axon-guidance genes in glutamatergic projection neurons, synaptic regulatory genes in both astrocytes and neurons, and pro-myelination factors in oligodendrocytes, suggesting widespread plasticity of cortical circuits. Indeed, whole-cell electrophysiology revealed that thyroid hormone alters excitatory and inhibitory synaptic transmission, an effect that requires thyroid hormone-induced gene regulatory programs in presynaptic neurons. Furthermore, thyroid hormone action in the frontal cortex regulates innate exploratory behaviors and causally promotes exploratory decision-making. Thus, thyroid hormone acts directly on the cerebral cortex in males to coordinate exploratory behaviors with whole-body metabolic state.
Keywords: body-brain coordination; exploration; metabolism; neuroscience; synaptic plasticity; thyroid hormone; transcriptionally regulated behavior.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests T.S.S. is an advisor and founder of Autobahn Therapeutics and is an inventor on several patents related to sobetirome and its derivatives. T.S.S. provided the sobetirome used in this study but was not involved in the study design, performing the work described, or in the analysis and presentation of the results.
Figures
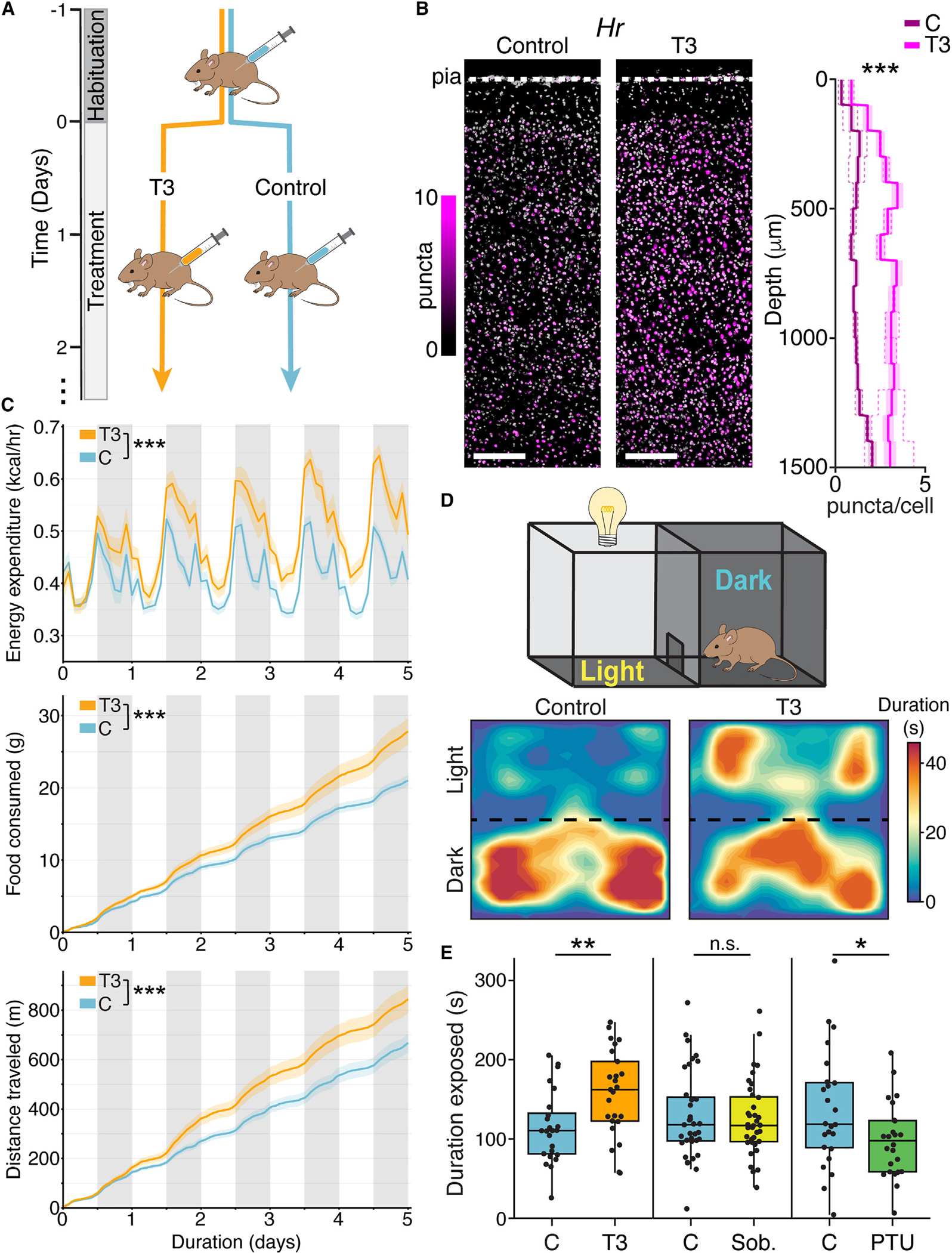
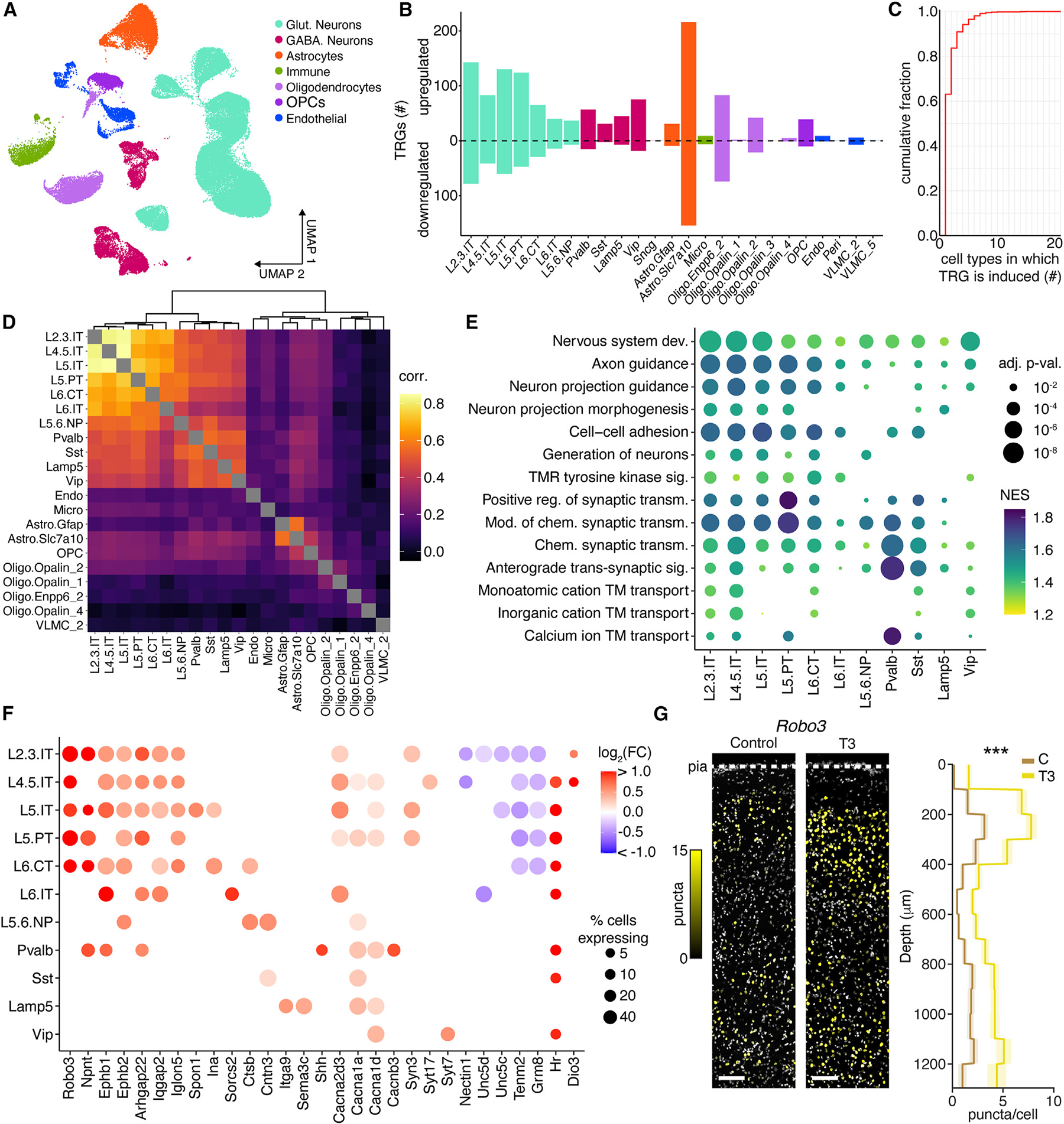
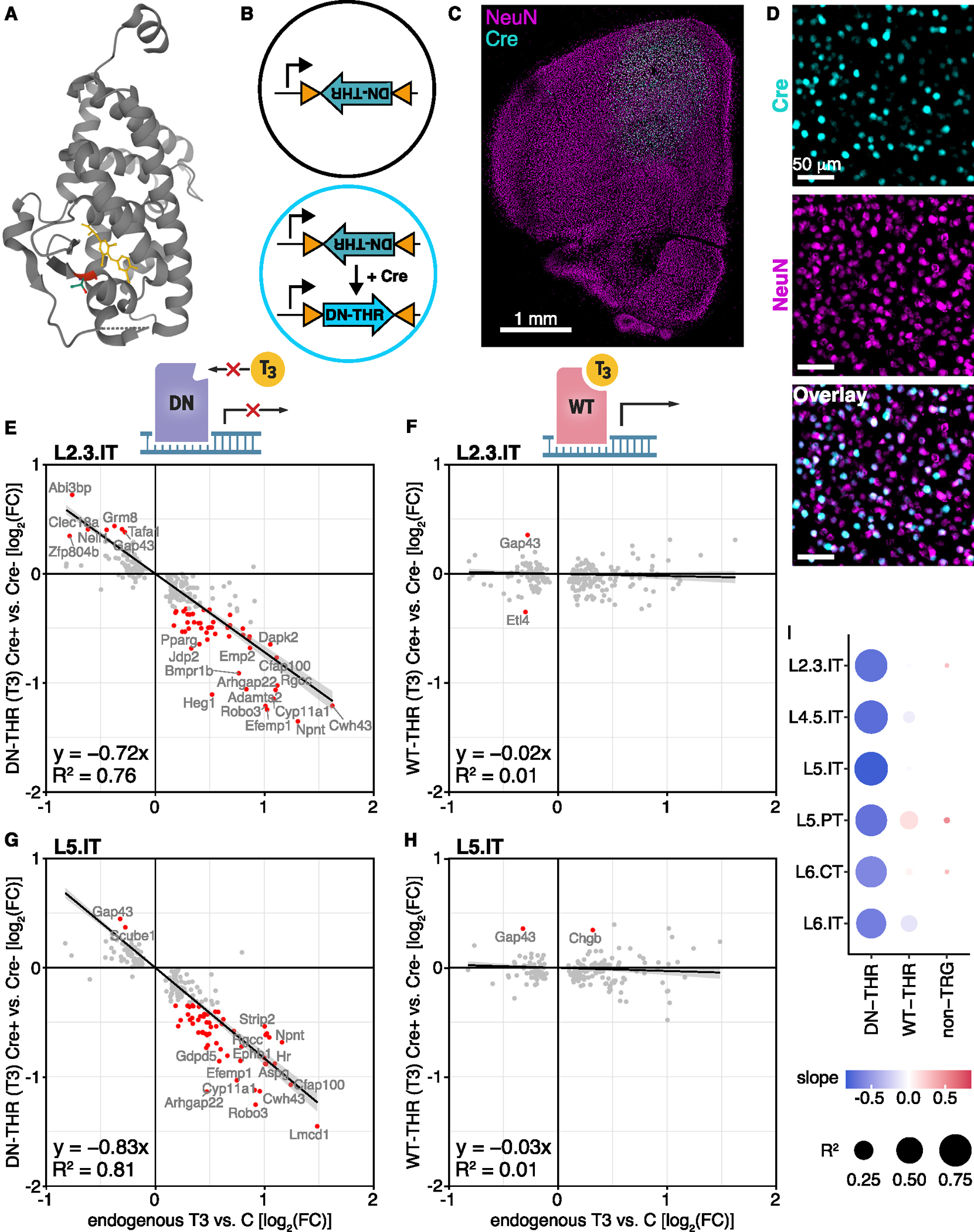
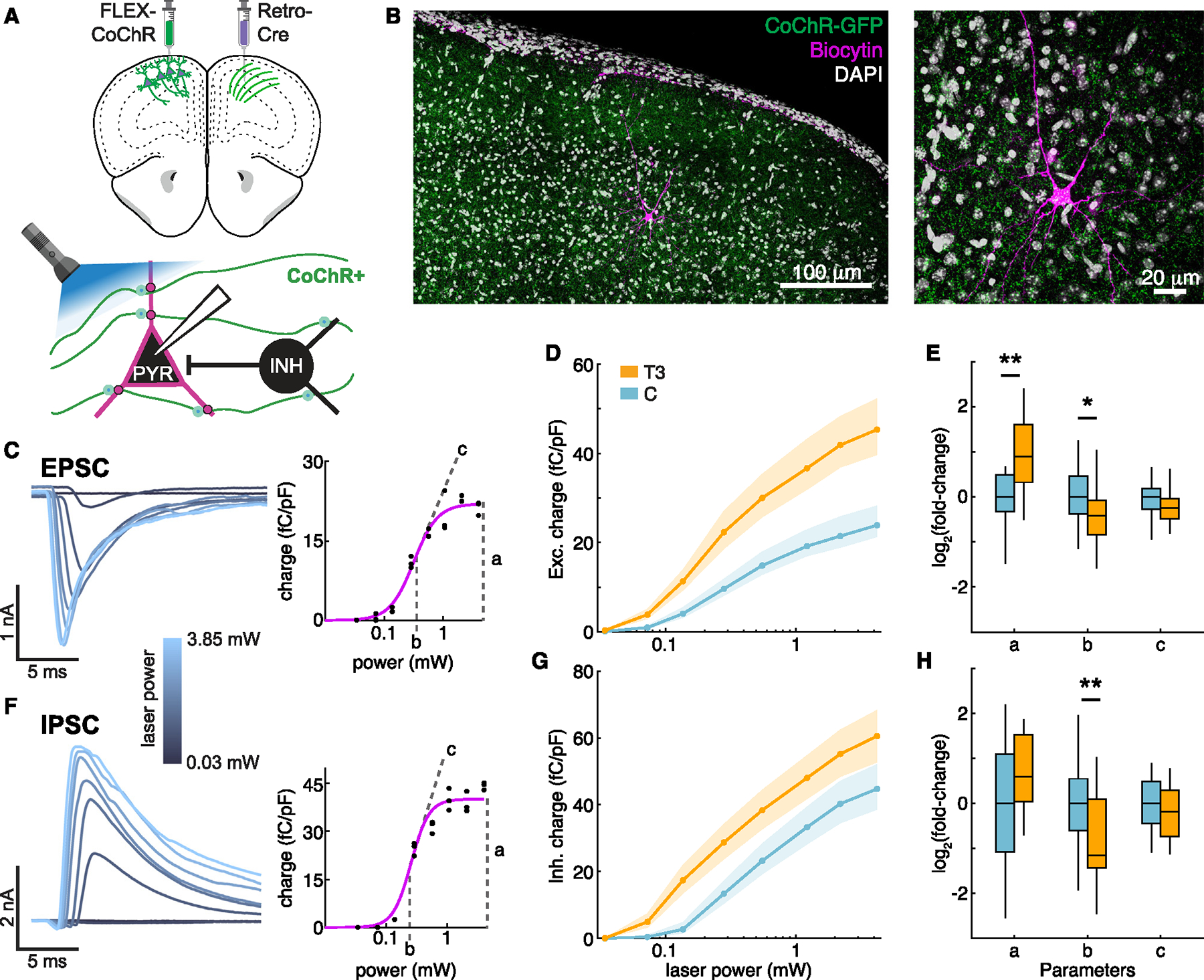
Update of
-
Thyroid hormone rewires cortical circuits to coordinate body-wide metabolism and exploratory drive.bioRxiv [Preprint]. 2023 Aug 10:2023.08.10.552874. doi: 10.1101/2023.08.10.552874. bioRxiv. 2023. Update in: Cell. 2024 Oct 3;187(20):5679-5697.e23. doi: 10.1016/j.cell.2024.07.041. PMID: 37609206 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
