

Complementary dual-virus strategy drives synthetic target and cognate T-cell engager expression for endogenous-antigen agnostic immunotherapy
- PMID: 39179564
- PMCID: PMC11343834
- DOI: 10.1038/s41467-024-51498-0
Complementary dual-virus strategy drives synthetic target and cognate T-cell engager expression for endogenous-antigen agnostic immunotherapy
Abstract
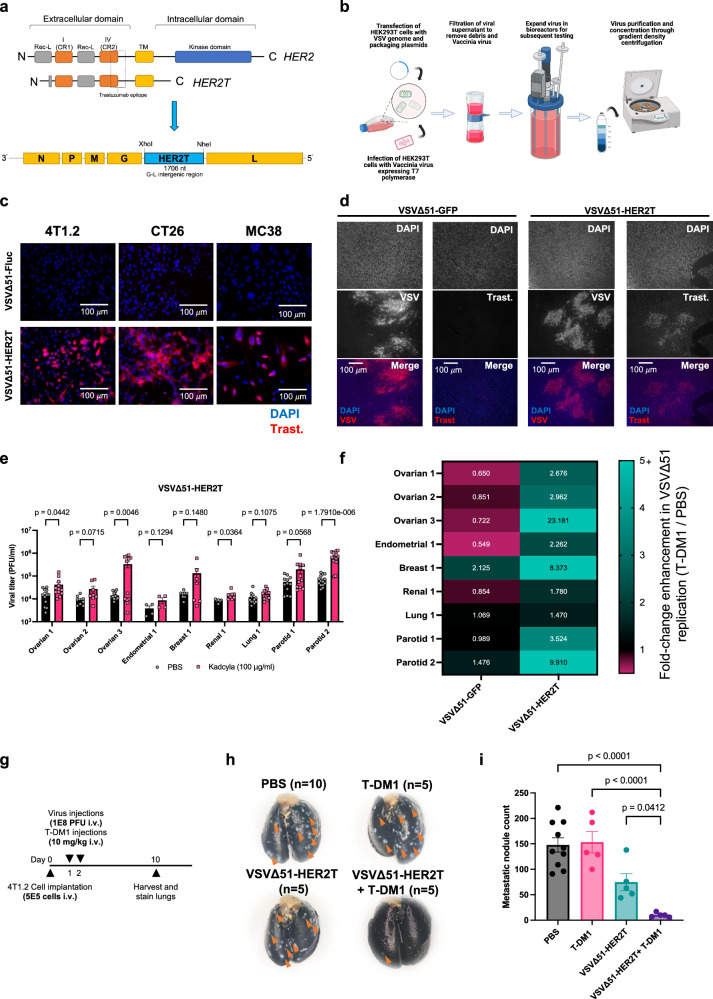
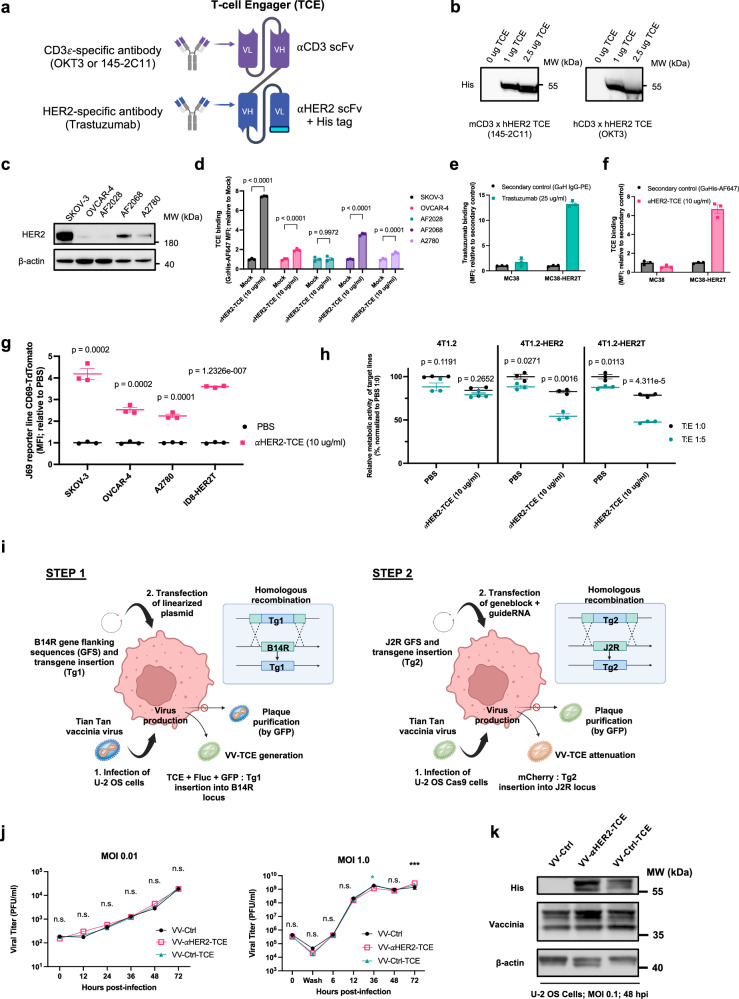
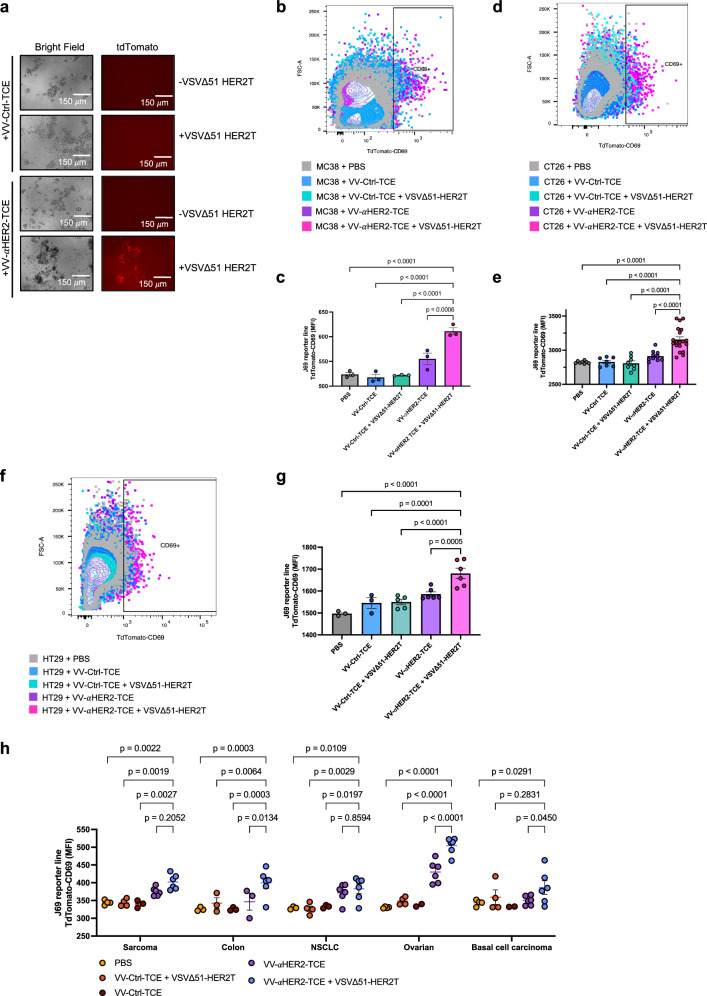
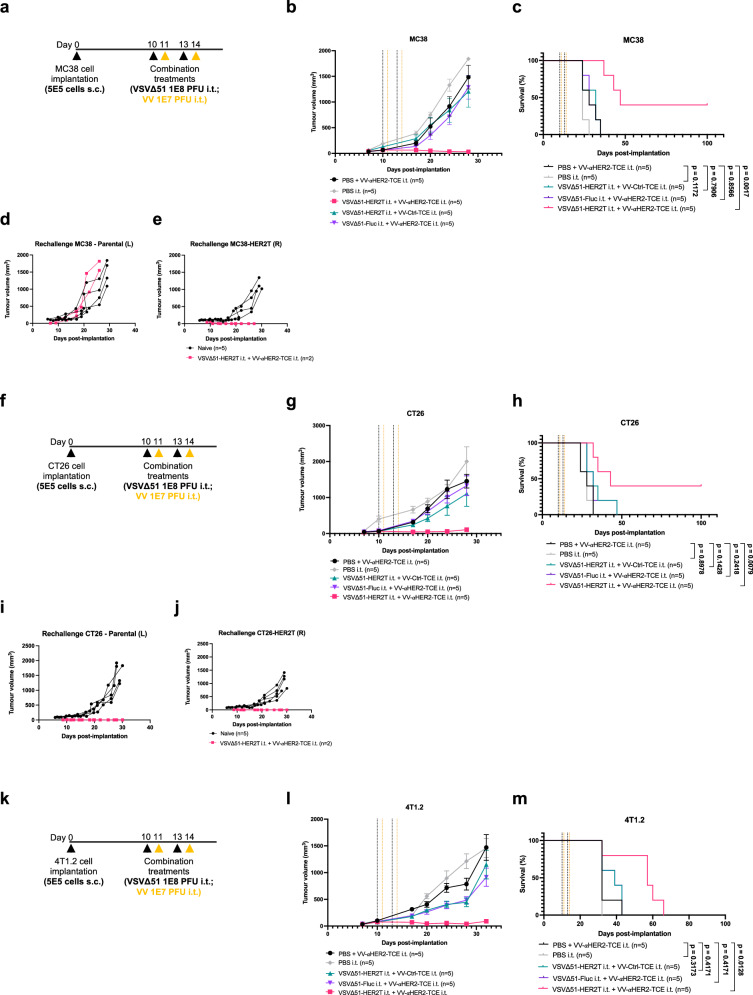
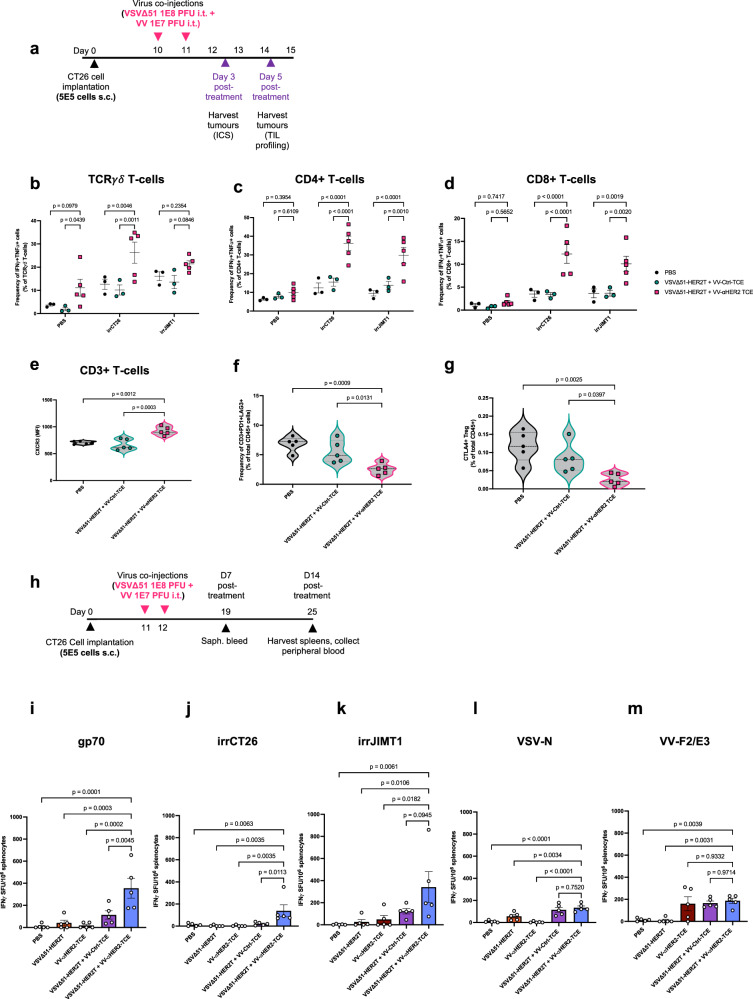
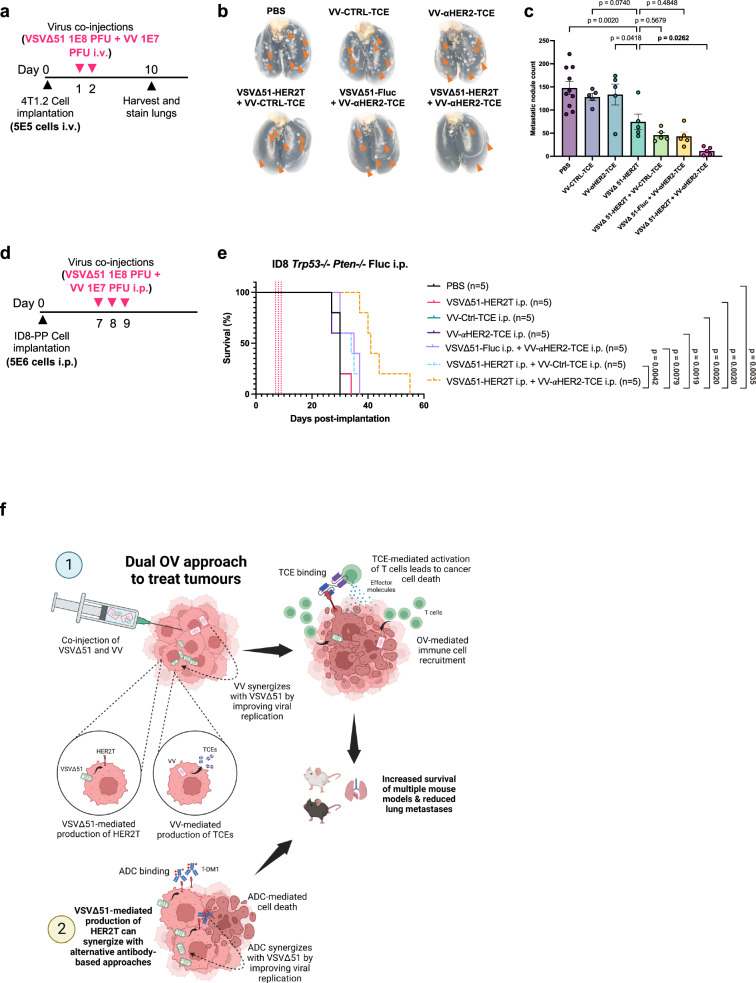
Targeted antineoplastic immunotherapies have achieved remarkable clinical outcomes. However, resistance to these therapies due to target absence or antigen shedding limits their efficacy and excludes tumours from candidacy. To address this limitation, here we engineer an oncolytic rhabdovirus, vesicular stomatitis virus (VSVΔ51), to express a truncated targeted antigen, which allows for HER2-targeting with trastuzumab. The truncated HER2 (HER2T) lacks signaling capabilities and is efficiently expressed on infected cell surfaces. VSVΔ51-mediated HER2T expression simulates HER2-positive status in tumours, enabling effective treatment with the antibody-drug conjugate trastuzumab emtansine in vitro, ex vivo, and in vivo. Additionally, we combine VSVΔ51-HER2T with an oncolytic vaccinia virus expressing a HER2-targeted T-cell engager. This dual-virus therapeutic strategy demonstrates potent curative efficacy in vivo in female mice using CD3+ infiltrate for anti-tumour immunity. Our findings showcase the ability to tailor the tumour microenvironment using oncolytic viruses, thereby enhancing compatibility with "off-the-shelf" targeted therapies.
© 2024. The Author(s).
Conflict of interest statement
Z.T., M.J.F.C, J.C.B., J-S.D. report having a pending patent directly related to this work entitled “Delivery vehicles expressing engineered antigens and uses thereof” (U.S. Provisional Patent Application No. 63/680, 672). The other authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
- 705952/Gouvernement du Canada | Instituts de Recherche en Santé du Canada | CIHR Skin Research Training Centre (Skin Research Training Centre)
- 377104/Gouvernement du Canada | Instituts de Recherche en Santé du Canada | CIHR Skin Research Training Centre (Skin Research Training Centre)
- 377104/Gouvernement du Canada | Instituts de Recherche en Santé du Canada | CIHR Skin Research Training Centre (Skin Research Training Centre)
- TFF-122868/Terry Fox Research Institute (Institut de Recherche Terry Fox)
- 708044/Canadian Cancer Society Research Institute (Société Canadienne du Cancer)
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
