

Regulation viral RNA transcription and replication by higher-order RNA structures within the nsp1 coding region of MERS coronavirus
- PMID: 39179600
- PMCID: PMC11343750
- DOI: 10.1038/s41598-024-70601-5
Regulation viral RNA transcription and replication by higher-order RNA structures within the nsp1 coding region of MERS coronavirus
Abstract
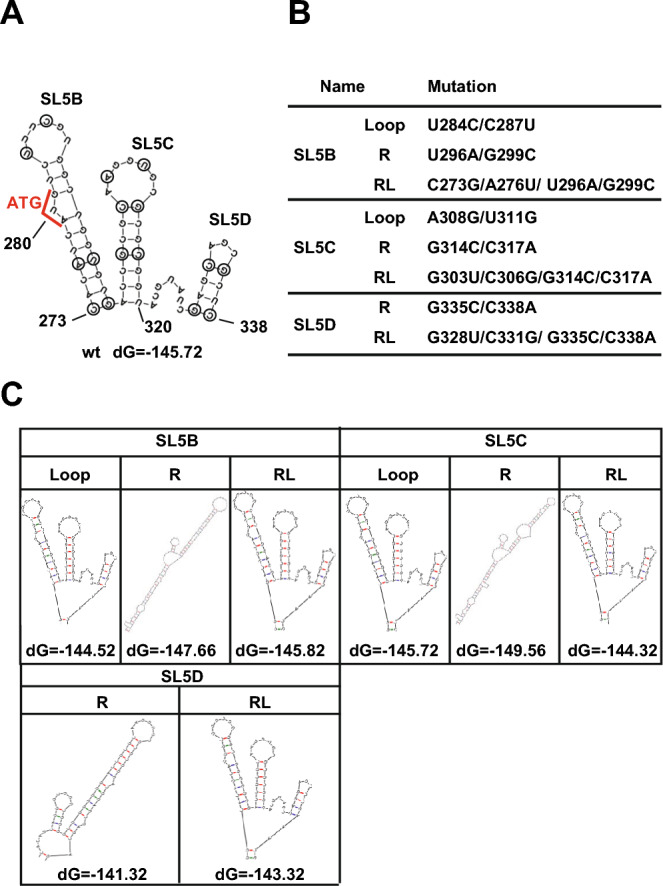
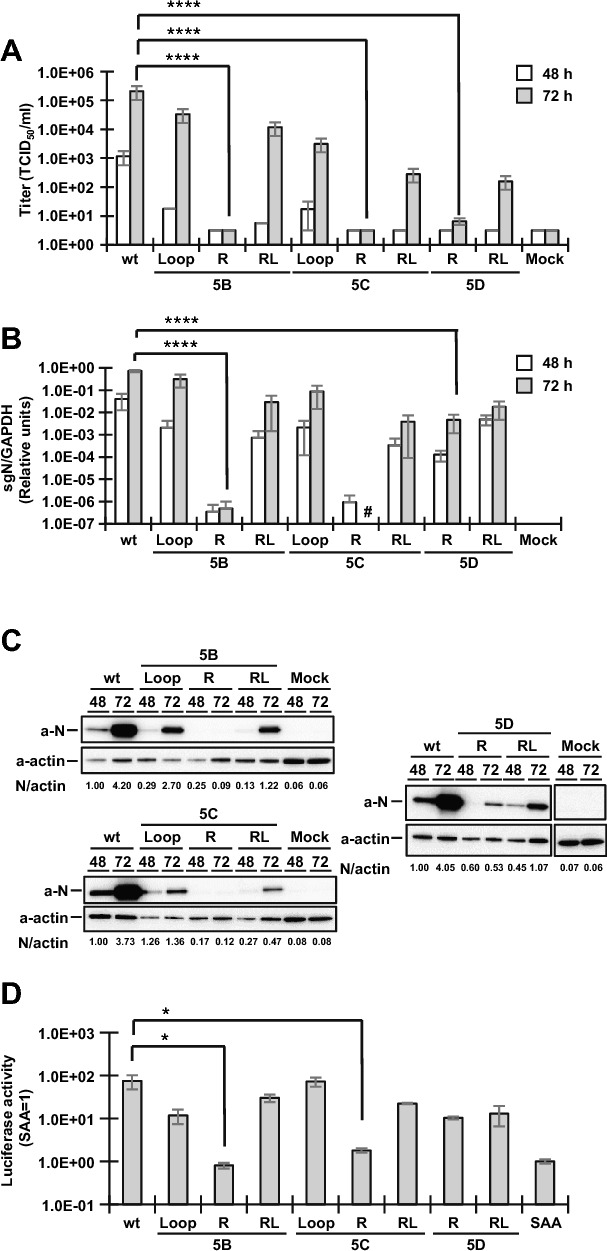
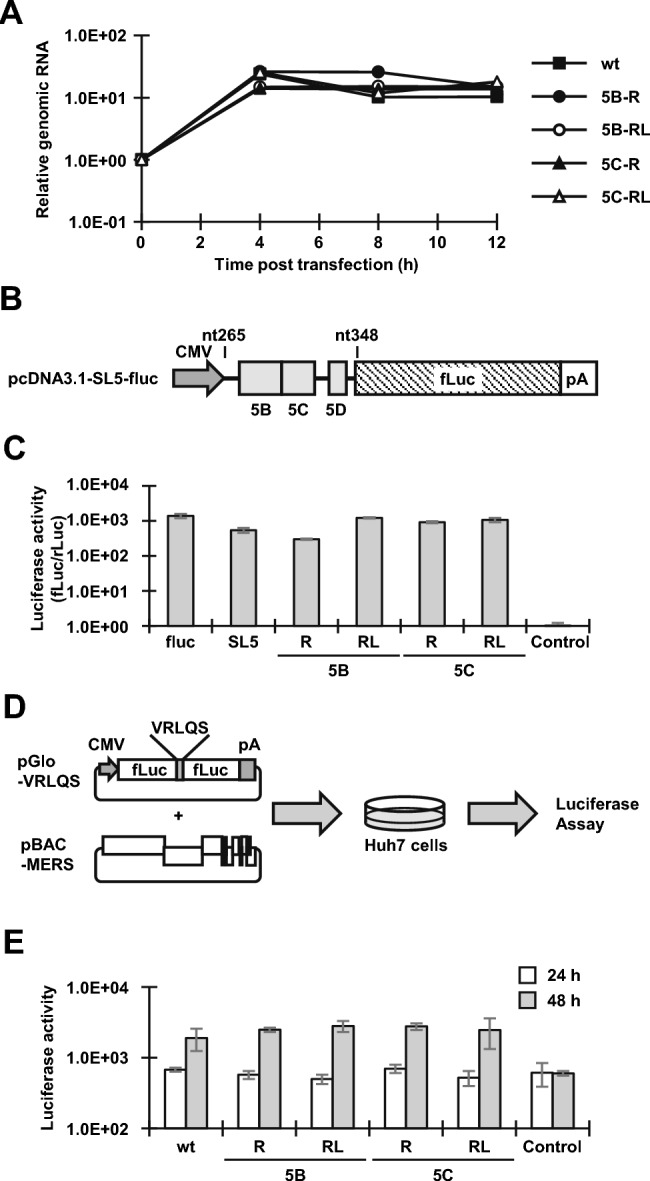
Coronavirus (CoV) possesses numerous functional cis-acting elements in its positive-strand genomic RNA. Although most of these RNA structures participate in viral replication, the functions of RNA structures in the genomic RNA of CoV in viral replication remain unclear. In this study, we investigated the functions of the higher-order RNA stem-loop (SL) structures SL5B, SL5C, and SL5D in the ORF1a coding region of Middle East respiratory syndrome coronavirus (MERS-CoV) in viral replication. Our approach, using reverse genetics of a bacterial artificial chromosome system, revealed that SL5B and SL5C play essential roles in the discontinuous transcription of MERS-CoV. In silico analyses predicted that SL5C interacts with a bulged stem-loop (BSL) in the 3' untranslated region, suggesting that the RNA structure of SL5C is important for viral RNA transcription. Conversely, SL5D did not affect transcription, but mediated the synthesis of positive-strand genomic RNA. Additionally, the RNA secondary structure of SL5 in the revertant virus of the SL5D mutant was similar to that of the wild-type, indicating that the RNA structure of SL5D can finely tune RNA replication in MERS-CoV. Our data indicate novel regulatory mechanisms of viral RNA transcription and replication by higher-order RNA structures in the MERS-CoV genomic RNA.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Expression and Cleavage of Middle East Respiratory Syndrome Coronavirus nsp3-4 Polyprotein Induce the Formation of Double-Membrane Vesicles That Mimic Those Associated with Coronaviral RNA Replication.mBio. 2017 Nov 21;8(6):e01658-17. doi: 10.1128/mBio.01658-17. mBio. 2017. PMID: 29162711 Free PMC article.
-
The Endonucleolytic RNA Cleavage Function of nsp1 of Middle East Respiratory Syndrome Coronavirus Promotes the Production of Infectious Virus Particles in Specific Human Cell Lines.J Virol. 2018 Oct 12;92(21):e01157-18. doi: 10.1128/JVI.01157-18. Print 2018 Nov 1. J Virol. 2018. PMID: 30111568 Free PMC article.
-
MERS coronavirus nsp1 participates in an efficient propagation through a specific interaction with viral RNA.Virology. 2017 Nov;511:95-105. doi: 10.1016/j.virol.2017.08.026. Epub 2017 Aug 23. Virology. 2017. PMID: 28843094 Free PMC article.
-
The structure and functions of coronavirus genomic 3' and 5' ends.Virus Res. 2015 Aug 3;206:120-33. doi: 10.1016/j.virusres.2015.02.025. Epub 2015 Feb 28. Virus Res. 2015. PMID: 25736566 Free PMC article. Review.
-
Functions of the 5'- and 3'-untranslated regions of tobamovirus RNA.Virus Res. 2015 Aug 3;206:82-9. doi: 10.1016/j.virusres.2015.01.028. Epub 2015 Feb 12. Virus Res. 2015. PMID: 25683511 Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
