

AQP1 differentially orchestrates endothelial cell senescence
- PMID: 39180980
- PMCID: PMC11388013
- DOI: 10.1016/j.redox.2024.103317
AQP1 differentially orchestrates endothelial cell senescence
Abstract
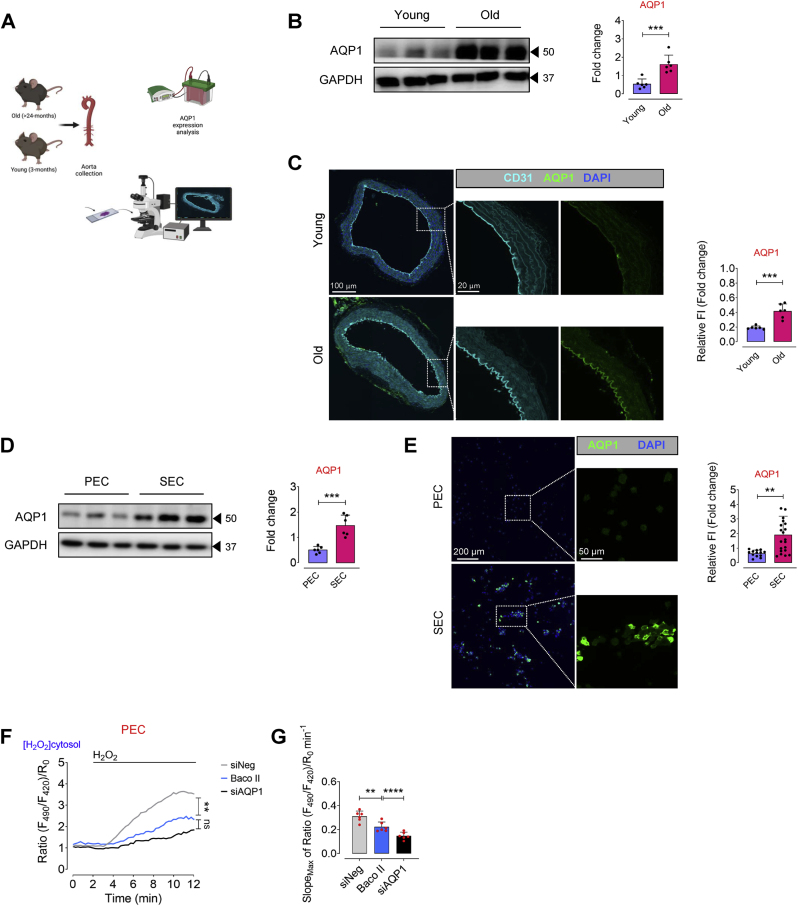
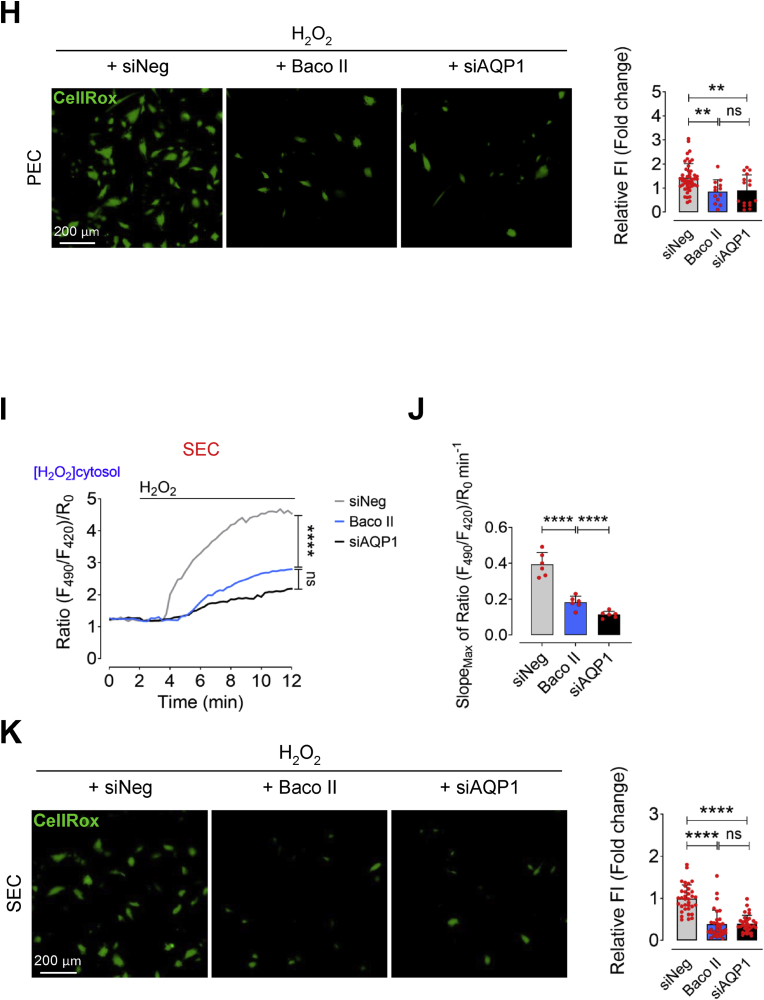
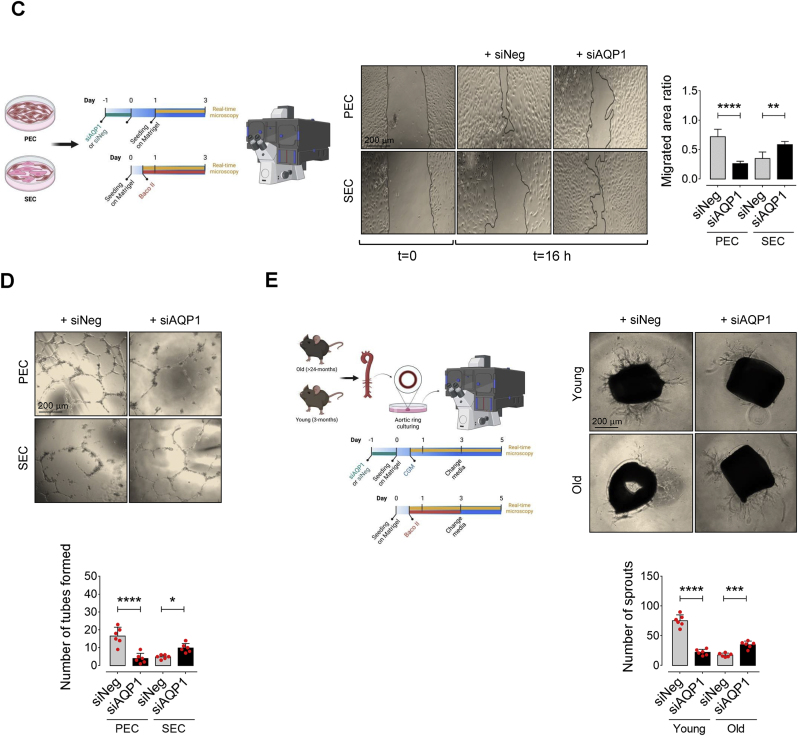
Accumulation of senescent endothelial cells (ECs) with age is a pivotal driver of cardiovascular diseases in aging. However, little is known about the mechanisms and signaling pathways that regulate EC senescence. In this report, we delineate a previously unrecognized role of aquaporin 1 (AQP1) in orchestrating extracellular hydrogen peroxide (H2O2)-induced cellular senescence in aortic ECs. Our findings underscore AQP1's differential impact on senescence hallmarks, including cell-cycle arrest, senescence-associated secretory phenotype (SASP), and DNA damage responses, intricately regulating angiogenesis. In proliferating ECs, AQP1 is crucial for maintaining angiogenic capacity, whereas disruption of AQP1 induces morphological and mitochondrial alterations, culminating in senescence and impaired angiogenesis. Conversely, Aqp1 knockdown or selective blockade of AQP1 in senescent ECs rescues the excess H2O2-induced cellular senescence phenotype and metabolic dysfunction, thereby ameliorating intrinsic angiogenic incompetence. Mechanistically, AQP1 facilitates H2O2 transmembrane transport, exacerbating oxidant-sensitive kinases CaMKII-AMPK. This process suppresses HDAC4 translocation, consequently de-repressing Mef2A-eNOS signaling in proliferating ECs. However, in senescent ECs, AQP1 overexpression is linked to preserved HDAC4-Mef2A complex and downregulation of eNOS signaling. Together, our studies identify AQP1 as a novel epigenetic regulator of HDAC4-Mef2A-dependent EC senescence and angiogenic potential, highlighting its potential as a therapeutic target for antagonizing age-related cardiovascular diseases.
Keywords: Aging; Angiogenesis; Aquaporin 1; Endothelial senescence; Epigenetic modification; Hydrogen peroxide.
Copyright © 2024 The Authors. Published by Elsevier B.V. All rights reserved.
Conflict of interest statement
Declaration of competing interest The authors declare the following financial interests/personal relationships which may be considered as potential competing interests: Soheil Saeedi reports was provided by University of Zurich. If there are other authors, they declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this paper.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
