Figure 4:. TEMPO imaging in anesthetized mice reveals traveling neocortical waves in the delta and gamma frequency bands that exhibit cross-frequency coupling.
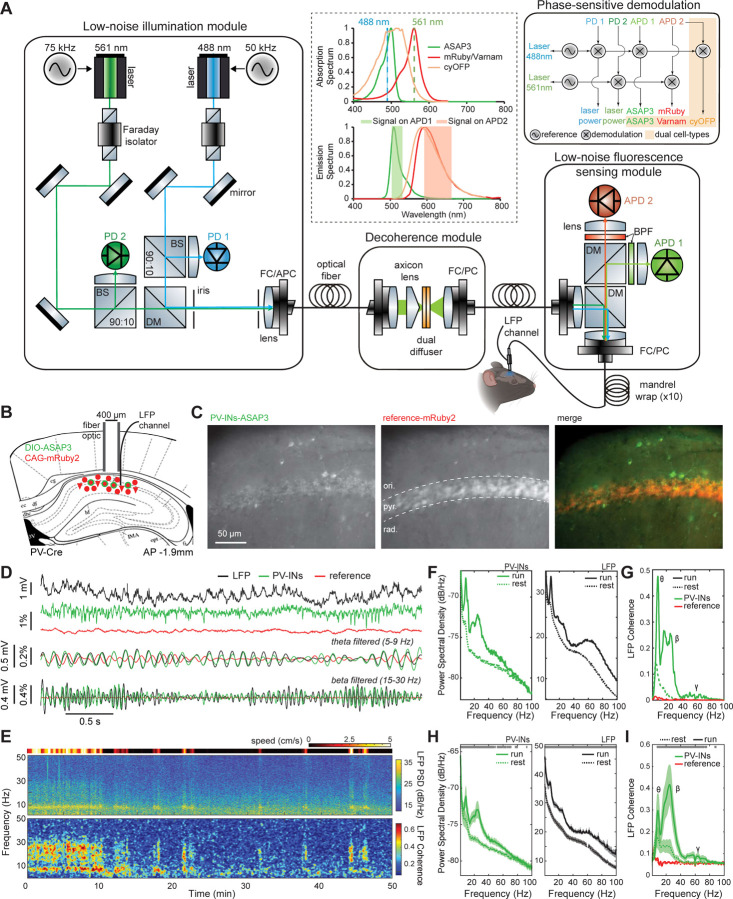
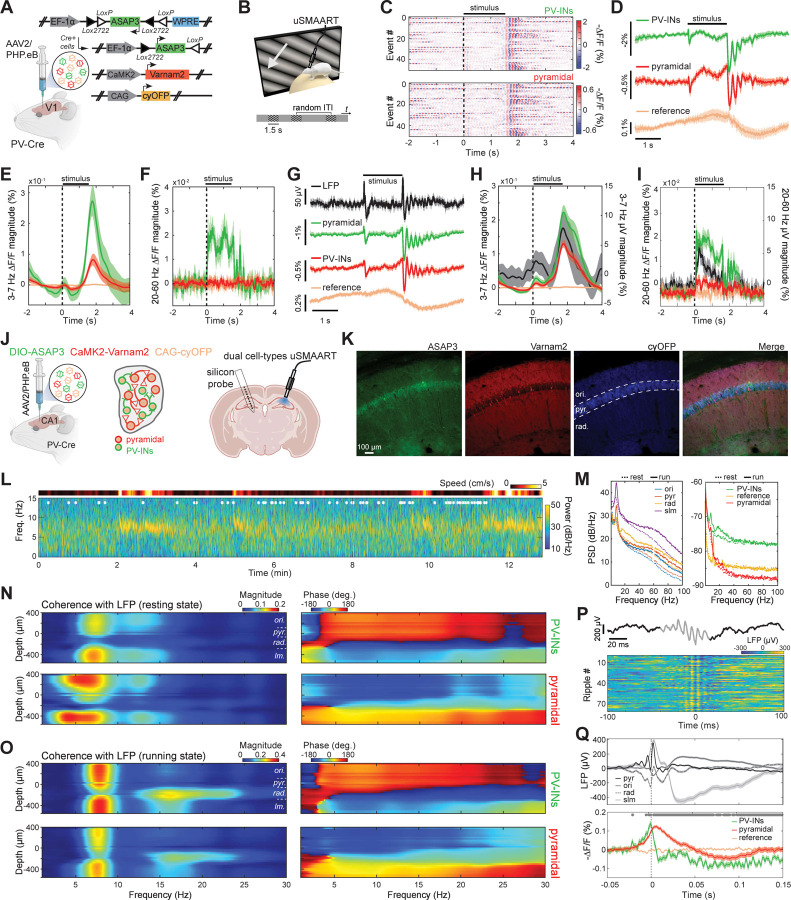
(A) Schematic of the TEMPO mesoscope. A pair of low-noise light-emitting diodes provides two-color illumination; a corresponding pair of photodiodes monitors their emission powers. The illumination reflects off a dual-band dichroic mirror (70 mm × 100 mm) and is focused onto the specimen by a 0.5 NA macro objective lens providing a 8-mm-diameter maximum field-of-view (FOV) when used in our system. Fluorescence returns through the objective lens and dual-band dichroic mirror. A short-pass dichroic mirror (70 mm × 100 mm) splits the fluorescence into two detection channels. After passing through a bandpass filter, fluorescence in each channel is focused onto a fast sCMOS camera by a tube lens (85 mm effective focal length). Inset: A view from the Allen Brain Atlas, showing the location of the glass cranial windows (dotted black circle; 7–8 mm diameter) used for Figures 4,5,7. Across the full area visible through the cranial window, the cameras acquired images at 130 Hz. In some studies, to increase the imaging speed to 300 Hz, we sampled a region-of-interest on the camera chip that covered the brain areas between the two black horizontal lines. Abbreviations: RSP (retrosplenial cortex), M1 (primary motor cortex), V1 (primary visual cortex), S1 (primary somatosensory cortex), BPF (bandpass filter), BS (beamsplitter), DM (dichroic mirror), LED (light-emitting diode), ND (neutral density), PD (photodiode). (B) Computer-assisted design mechanical drawing of the mesoscope. Lower inset, magnified view of the large custom fluorescence filter set. Upper inset: Timing protocol for voltage imaging using a single green GEVI plus the mRuby2 reference fluor. Illumination from both LEDs is continuous (bottom two traces). Image acquisition by the two cameras is initiated by an external trigger, ensuring that the image pairs are temporally aligned (top two traces; 800 Mbytes · s–1 data rate per camera). (C) Top, We retro-orbitally co-injected a pair of AAV2/PHP.eB viruses to co-express red fluorescent mRuby2 and a green fluorescent GEVI, ASAP3. Bottom, One virus expresses mRuby2 via the CAG promoter; the other allows Cre-dependent expression of ASAP3 via the EF-1α promoter. By using Cux2-CreERT2 or PV-Cre mice, we performed voltage-imaging studies of neocortical layer 2/3 pyramidal (L2/3) or PV cells, respectively. (D) Two examples of traveling voltage waves in the delta frequency band, shown in ASAP3 image sequences (50 ms between images) taken at 130 Hz in ketamine-xylazine-anesthetized Cux2-CreERT2 mice. Images underwent unmixing (Figure S5) to remove hemodynamic changes captured in the mRuby2 channel but were otherwise unfiltered. Brain area boundaries (see (A) inset), are superposed on the last image in each sequence. In each case, a depolarization (denoted by red hues) sweeps across cortex in the anterior to posterior (A-P) direction. Data in (E–R) are also from ketamine-xylazine-anesthetized mice and were acquired at 130 Hz in (D–J) and 300 Hz in (K–R). (E) Color plot (top) showing the anterior to posterior propagation of the two traveling waves in (D). At each time point (x-axis) and for each A-P coordinate (y-axis), we averaged fluorescence values along the medio-lateral direction. Arrows in 3 different shades of green mark 3 different positions along the A-P axis for which voltage-dependent fluorescence traces are plotted (bottom) in corresponding colors. (F) Flow maps showing local propagation directions of voltage depolarization for a pair of individual delta waves observed in example Cux2-CreERT2 (top) and PV-Cre (bottom) mice. Flow vectors are all normalized to have the same length. (G) Distributions of delta wave propagation speed across all delta events seen in two Cux2-CreERT2 (top) and two (PV-Cre) (bottom) mice, computed near the center of area V1 (marked by black dots in (I)), where there was consistent anterior to posterior propagation. Insets: Polar histograms showing distributions of wave propagation direction for the same 4 mice at the center of V1, revealing the approximate alignment of wave propagation with the A-P axis in all 4 mice (n = 313, 320, 405 and 200 delta events in the individual mice). (H) During the peaks of the delta waves, we found enhanced activity in the gamma (30–60 Hz) frequency band. In two example Cux2-CreERT2 (top) and PV-Cre mice (bottom), the amplitudes of gamma oscillations increased during delta wave depolarizations up to ~4-fold over baseline values in brain areas V1 and RSP and to a lesser extent in other areas. (I) Maps of peak correlation coefficients, r, for the same mice as in (H), computed for each spatial point by calculating the temporal correlation function between the local fluorescence trace and that at the center of V1 (black dots) and then finding this function’s maximum value. (J) Maps of peak correlation coefficients, computed as in (I) but using gamma bandpass-filtered fluorescence traces, show that the gamma oscillations were less spatially coherent than the delta waves. (K) To study gamma activity in greater detail, we acquired a subset of the video data at 300 Hz (K–R) over a more limited FOV (see inset of (A)). Top: Example fluorescence trace of voltage activity at the center of V1 in the same Cux2-CreERT2 mouse as in (H-J), showing ongoing delta waves. Bottom: Gamma-band filtered (35–100 Hz) version of the top trace, revealing increases in gamma-band activity during the peaks of the delta oscillations. (L) Top: Example fluorescence traces of voltage activity at the center of V1, from each of the two mice in (H-J), temporally aligned to the peak of delta wave depolarization to reveal the consistent waveform of delta activity (n = 260 waves shown per mouse). Bottom: Gamma-band filtered (35–100 Hz) versions of the same traces reveal delta-gamma coupling as in (K). (M) Mean time-dependent amplitudes of gamma-band (35–100 Hz) activity in the voltage (solid lines) and reference (dashed lines) signals, for each of the 4 mice in (G), computed by averaging over all delta events in each mouse, identified as in (L) and aligned to the peak of the delta oscillation using the wavelet spectrogram. Voltage but not reference channel signals showed increased gamma band activity during the depolarization phase of delta oscillations. Shading: 95% C.I. (N) Top: Magnified views of two example delta depolarization events at the center of V1 in the same mouse as in (K). Middle: Gamma-filtered (35–100 Hz) versions of the same traces, highlighting gamma events near the end of each delta depolarization. Bottom: Gamma-filtered traces of activity during Event 1, from the color-corresponding locations marked with green-shaded dots in the upper rightmost image of panel (O). (O) Sequences of gamma-band (35–100 Hz) filtered images (3 ms between successive frames), showing the spatiotemporal dynamics in V1 of the same two delta depolarization events as in (N). Green dots mark the anatomic locations for the voltage traces in the bottom panel of (N). For display purposes only, the images shown were spatially low-pass filtered using a Gaussian filter (156 μm FWHM). (P, Q) Left panels: Flow maps showing the local wave propagation directions during the same two individual gamma wave events as in (N). As in (F), all flow vectors are normalized to have the same length. Right panels: Histograms showing the distributions of propagation speed across the brain region shown in (O), for the same two gamma events as in the left panels. Unlike delta waves, which traveled along the A-P axis, the gamma waves illustrated here traveled more aligned to the medio-lateral (M-L) axis and had much faster speeds than the accompanying delta waves (compare to (G)). Insets: Polar histograms showing the distributions of propagation direction for the two gamma events, computed across the flow maps of the left panels. (R) Histograms of propagation speed, aggregated across n = 20 events and all spatial bins (62.5 µm wide) in V1, in the same two mice as in (H-J) (n = 200 and 300 waves, respectively, in the Cux2-CreERT2 and PV-Cre mice). Insets: Polar histograms of the directions of gamma wave propagation, showing that in both L2/3 pyramidal and PV cells the gamma waves traveled approximately in the M-L direction, roughly orthogonal to the propagation directions of their carrier delta waves (E–G).