

Animal and bacterial viruses share conserved mechanisms of immune evasion
- PMID: 39197447
- PMCID: PMC11455605
- DOI: 10.1016/j.cell.2024.07.057
Animal and bacterial viruses share conserved mechanisms of immune evasion
Abstract
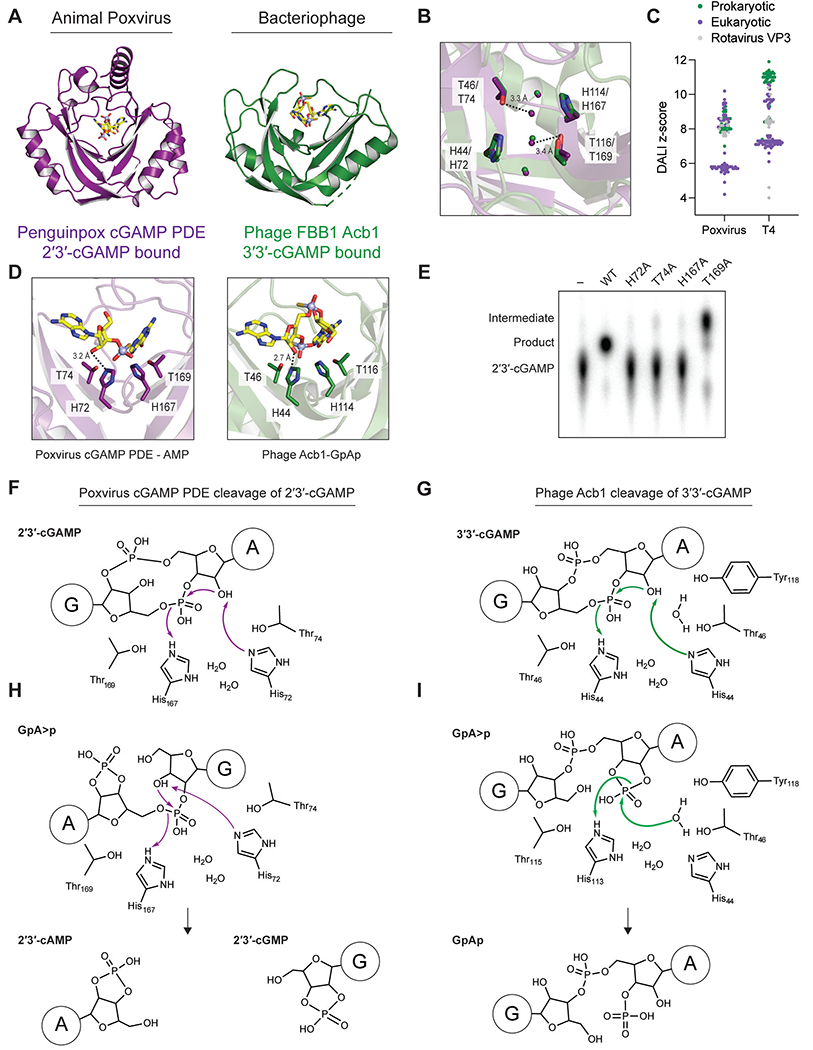
Animal and bacterial cells sense and defend against viral infections using evolutionarily conserved antiviral signaling pathways. Here, we show that viruses overcome host signaling using mechanisms of immune evasion that are directly shared across the eukaryotic and prokaryotic kingdoms of life. Structures of animal poxvirus proteins that inhibit host cGAS-STING signaling demonstrate architectural and catalytic active-site homology shared with bacteriophage Acb1 proteins, which inactivate CBASS anti-phage defense. In bacteria, phage Acb1 proteins are viral enzymes that degrade host cyclic nucleotide immune signals. Structural comparisons of poxvirus protein-2'3'-cGAMP and phage Acb1-3'3'-cGAMP complexes reveal a universal mechanism of host nucleotide immune signal degradation and explain kingdom-specific additions that enable viral adaptation. Chimeric bacteriophages confirm that animal poxvirus proteins are sufficient to evade immune signaling in bacteria. Our findings identify a mechanism of immune evasion conserved between animal and bacterial viruses and define shared rules that explain host-virus interactions across multiple kingdoms of life.
Keywords: CBASS; antiviral immunity; cGAS; cGLR; cyclic nucleotide; immune evasion; poxvirus.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests J.A.D. is a co-founder of Caribou Biosciences, Editas Medicine, Scribe Therapeutics, Intellia Therapeutics, and Mammoth Biosciences. J.A.D. is a scientific advisory board member of Vertex, Caribou Biosciences, Intellia Therapeutics, Scribe Therapeutics, Mammoth Biosciences, Algen Biotechnologies, Felix Biosciences, The Column Group, and Inari Agriculture. J.A.D. is Chief Science Advisor to Sixth Street, a Director at Johnson & Johnson, and Altos and Tempus and has research projects sponsored by Apple Tree Partners and Roche.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
