

B-cell intrinsic RANK signaling cooperates with TCL1 to induce lineage-dependent B-cell transformation
- PMID: 39198400
- PMCID: PMC11358282
- DOI: 10.1038/s41408-024-01123-6
B-cell intrinsic RANK signaling cooperates with TCL1 to induce lineage-dependent B-cell transformation
Abstract
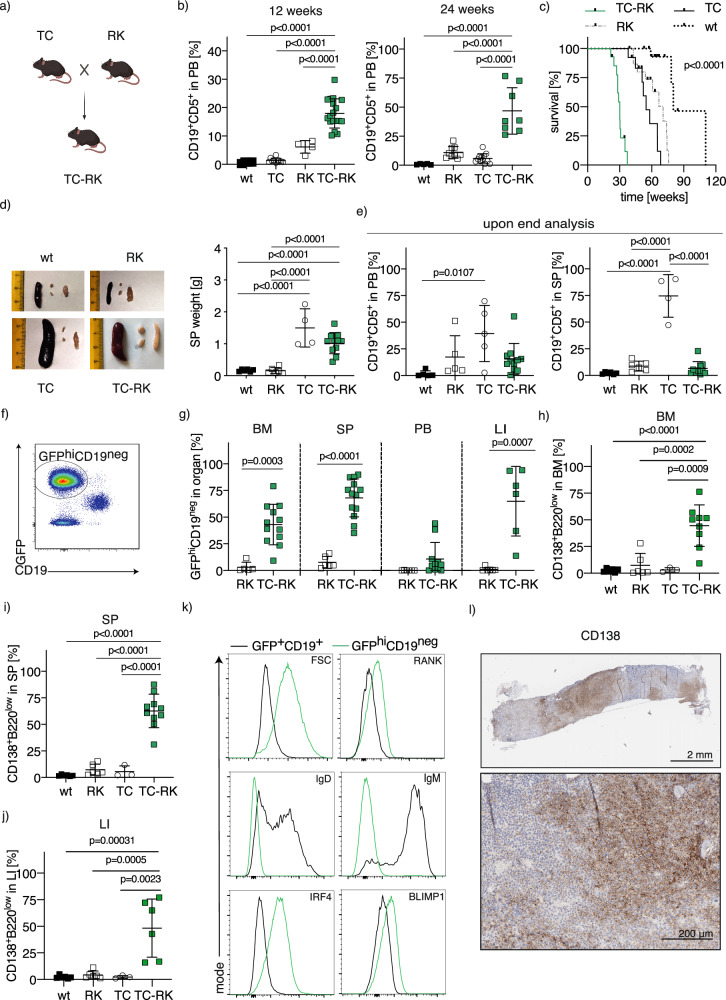
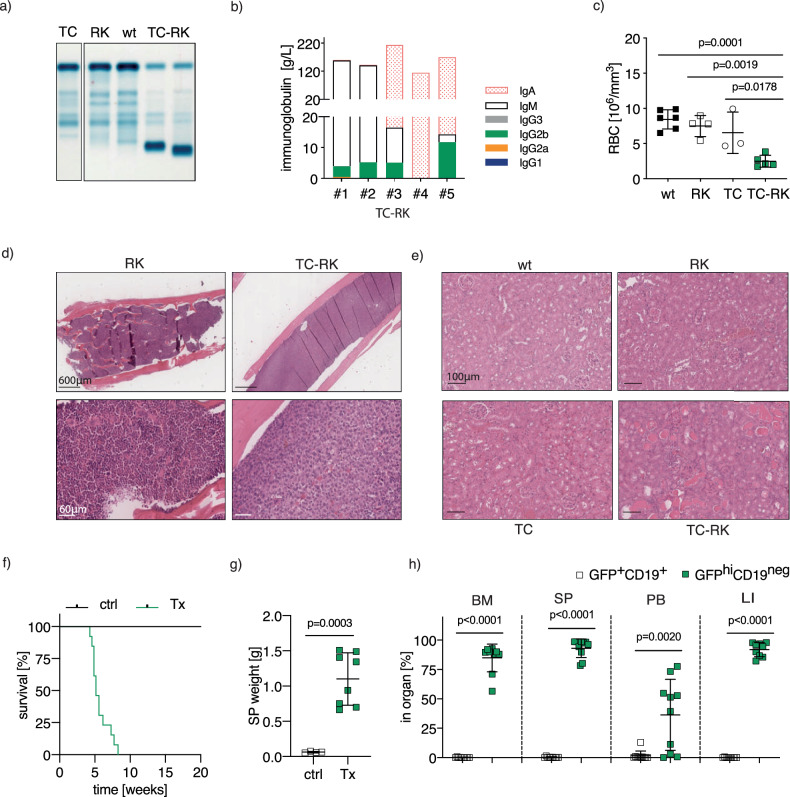
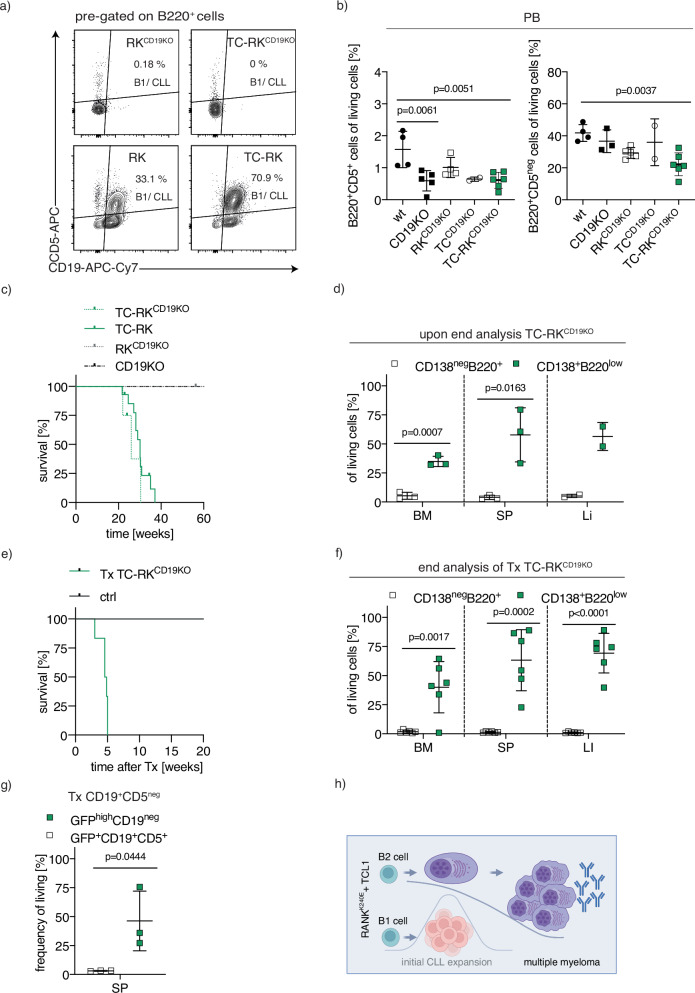
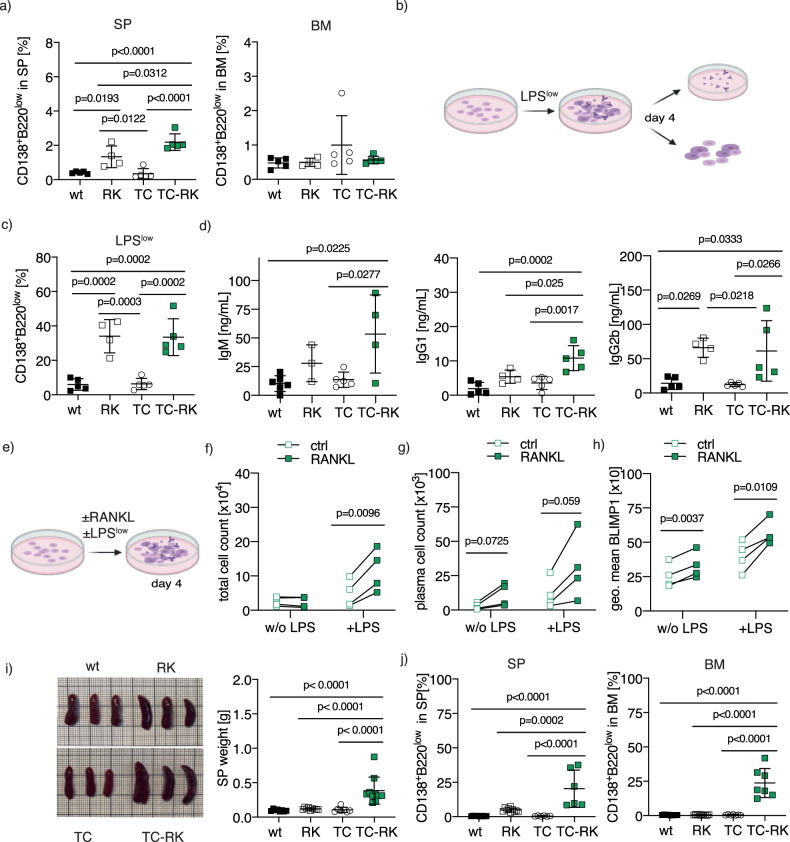
B-cell malignancies, such as chronic lymphocytic leukemia (CLL) and multiple myeloma (MM), remain incurable, with MM particularly prone to relapse. Our study introduces a novel mouse model with active RANK signaling and the TCL1 oncogene, displaying both CLL and MM phenotypes. In younger mice, TCL1 and RANK expression expands CLL-like B1-lymphocytes, while MM originates from B2-cells, becoming predominant in later stages and leading to severe disease progression and mortality. The induced MM mimics human disease, exhibiting features like clonal plasma cell expansion, paraproteinemia, anemia, and kidney and bone failure, as well as critical immunosurveillance strategies that promote a tumor-supportive microenvironment. This research elucidates the differential impacts of RANK activation in B1- and B2-cells and underscores the distinct roles of single versus combined oncogenes in B-cell malignancies. We also demonstrate that human MM cells express RANK and that inhibiting RANK signaling can reduce MM progression in a xenotransplantation model. Our study provides a rationale for further investigating the effects of RANK signaling in B-cell transformation and the shaping of a tumor-promoting microenvironment.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
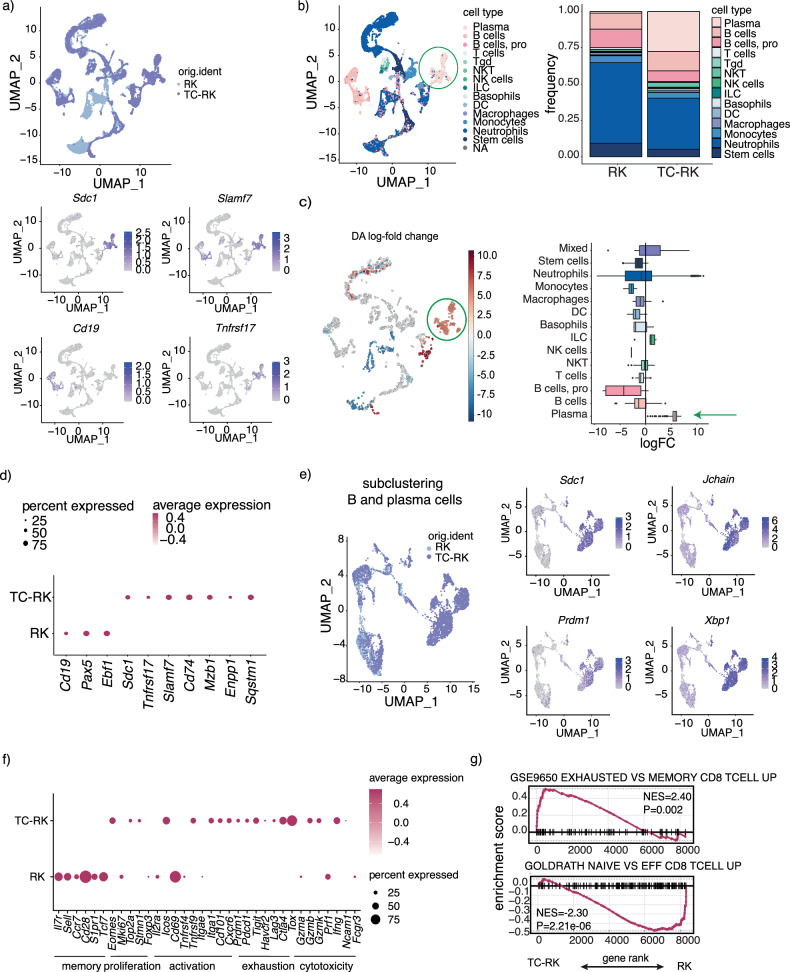
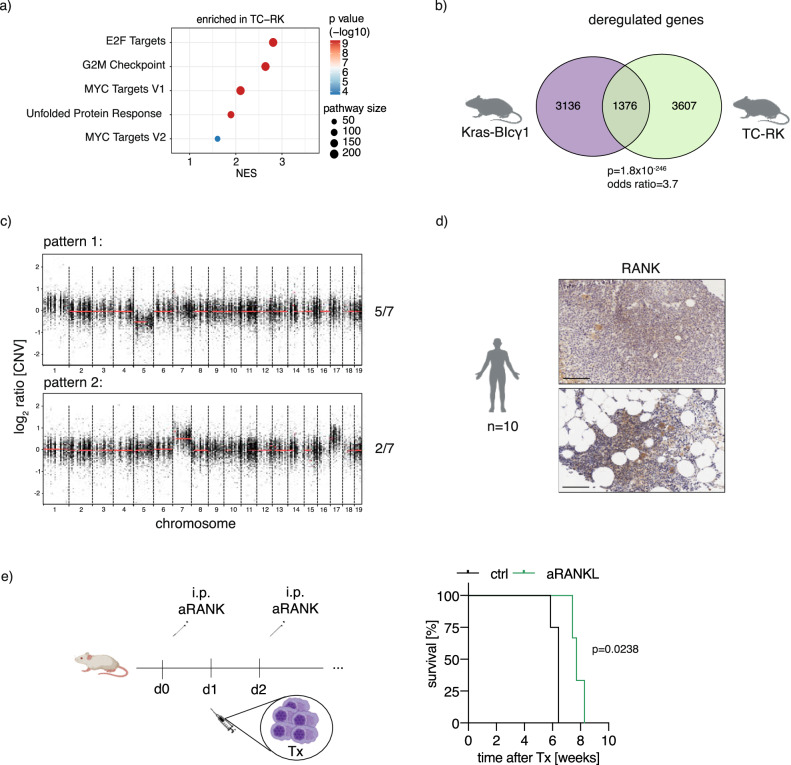
References
Publication types
MeSH terms
Substances
Grants and funding
- 516923543/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 505372148/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 2851/6-1/Deutsche Forschungsgemeinschaft (German Research Foundation)
- BA 2851/7-1/Deutsche Forschungsgemeinschaft (German Research Foundation)
- 70114720/Deutsche Krebshilfe (German Cancer Aid)
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
