

Sugar assimilation underlying dietary evolution of Neotropical bats
- PMID: 39198571
- PMCID: PMC11383804
- DOI: 10.1038/s41559-024-02485-7
Sugar assimilation underlying dietary evolution of Neotropical bats
Abstract
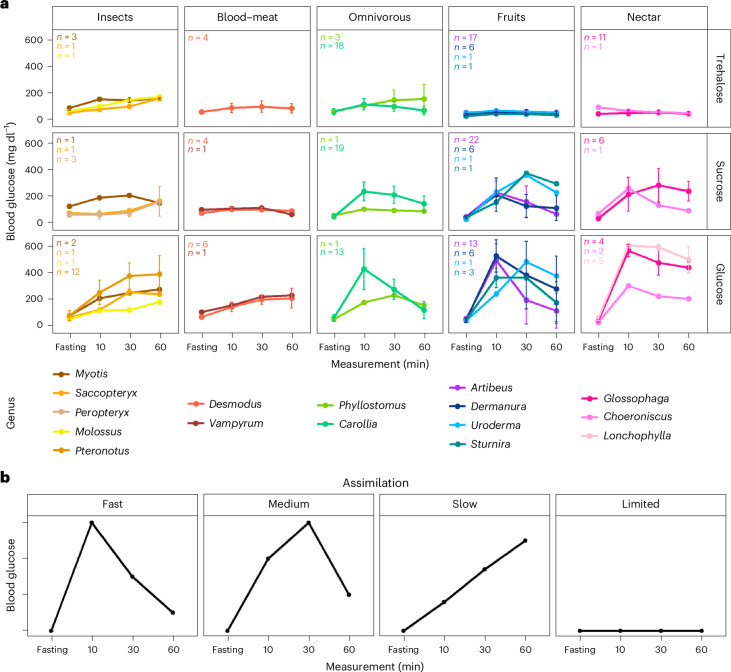
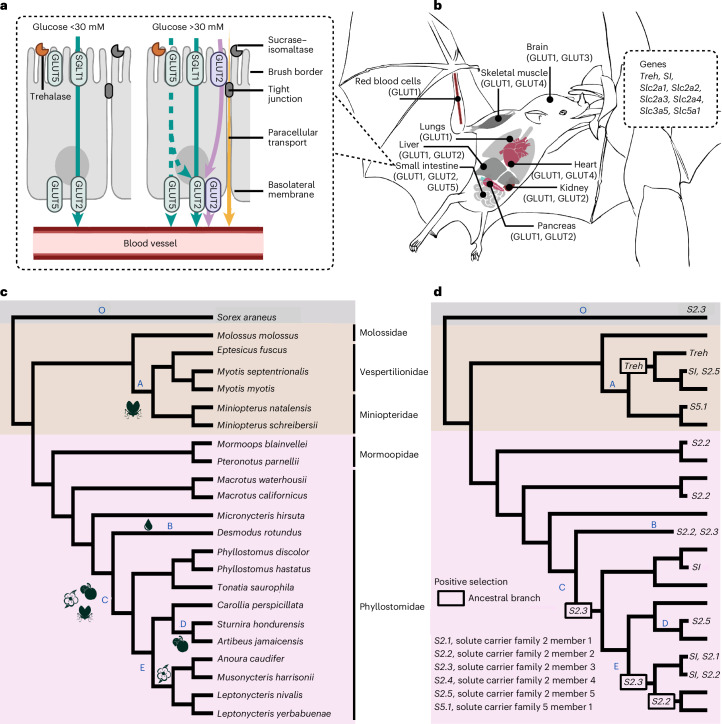
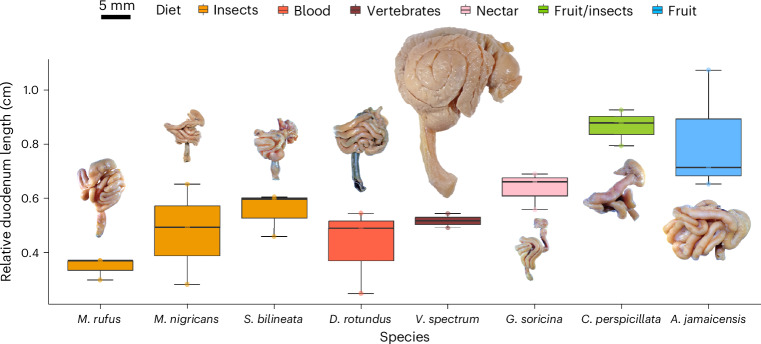
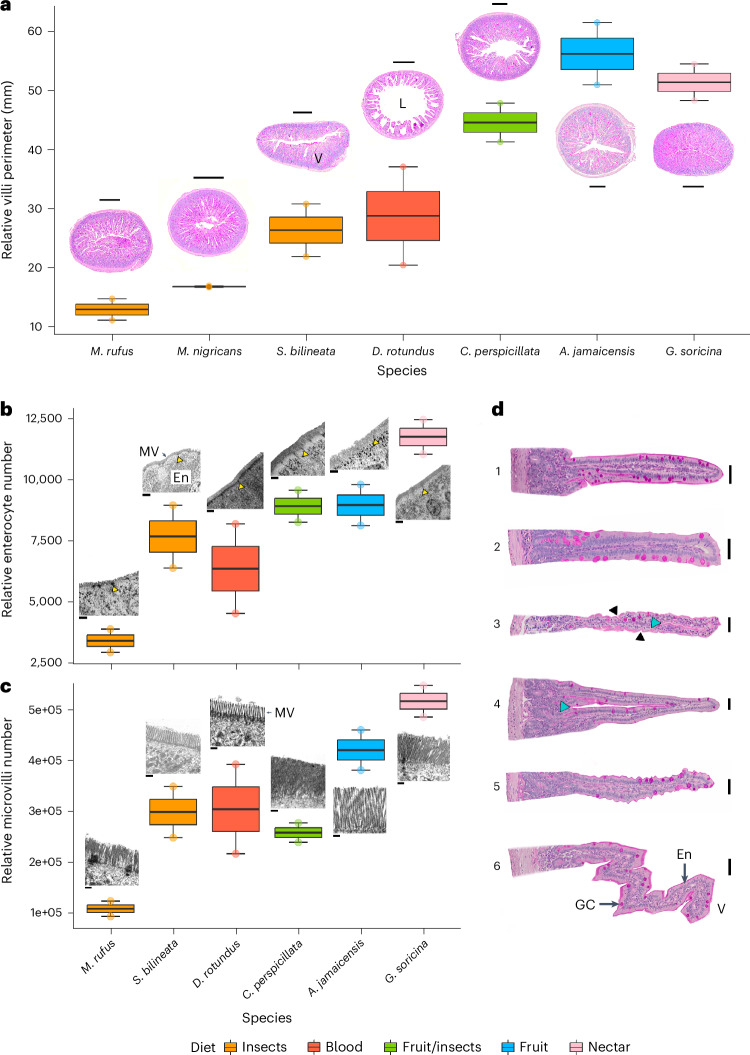
Dietary specializations in animals lead to adaptations in morphology, anatomy and physiology. Neotropical bats, with their high taxonomic and trophic diversity, offer a unique perspective on diet-driven evolutionary adaptations. Here we assess the metabolic response to different dietary sugars among wild-caught bats. We found that insectivorous bats had a pronounced metabolic response to trehalose, whereas bats with nectar and fruit-based diets showed significantly higher blood glucose levels in response to glucose and sucrose, reaching levels over 750 mg dl-1. The genomic analysis of 22 focal species and two outgroup species identified positive selection for the digestive enzyme trehalase in insect eaters, while sucrase-isomaltase showed selection in lineages with omnivorous and nectar diets. By examining anatomical and cellular features of the small intestine, we discovered that dietary sugar proportion strongly impacted numerous digestive traits, providing valuable insight into the physiological implications of molecular adaptations. Using hybridization chain reaction (HCR) RNA fluorescence in situ hybridization, we observed unusually high expression in the glucose transporter gene Slc2a2 in nectar bats, while fruit bats increased levels of Slc5a1 and Slc2a5. Overall, this study highlights the intricate interplay between molecular, morphological and physiological aspects of diet evolution, offering new insights into the mechanisms of dietary diversification and sugar assimilation in mammals.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures







References
-
- Pauli, J. N., Peery, M. Z., Fountain, E. D. & Karasov, W. H. Arboreal folivores limit their energetic output, all the way to slothfulness. Am. Nat.188, 196–204 (2016). - PubMed
-
- Camacho, J. et al. Peramorphosis, an evolutionary developmental mechanism in Neotropical bat skull diversity. Dev. Dyn.248, 1129–1143 (2019). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
