

Spatially clustered type I interferon responses at injury borderzones
- PMID: 39198639
- PMCID: PMC11374671
- DOI: 10.1038/s41586-024-07806-1
Spatially clustered type I interferon responses at injury borderzones
Abstract
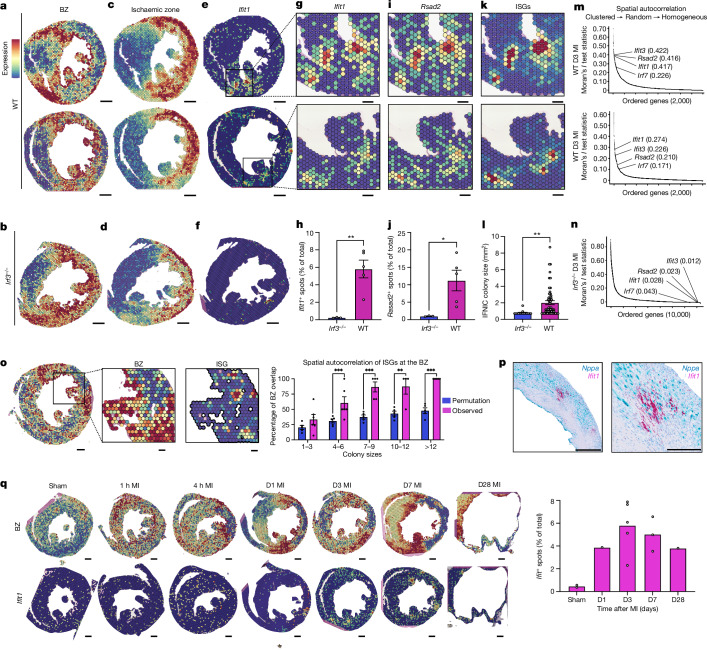
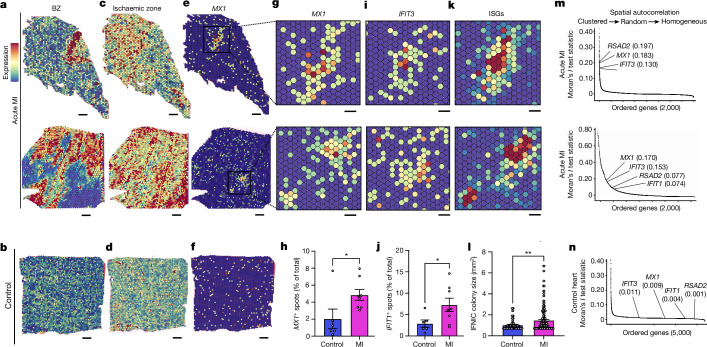
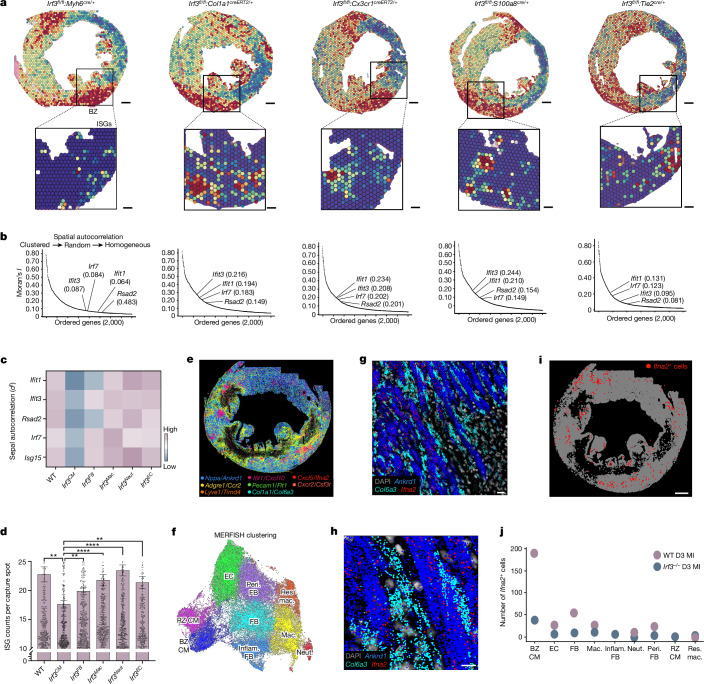
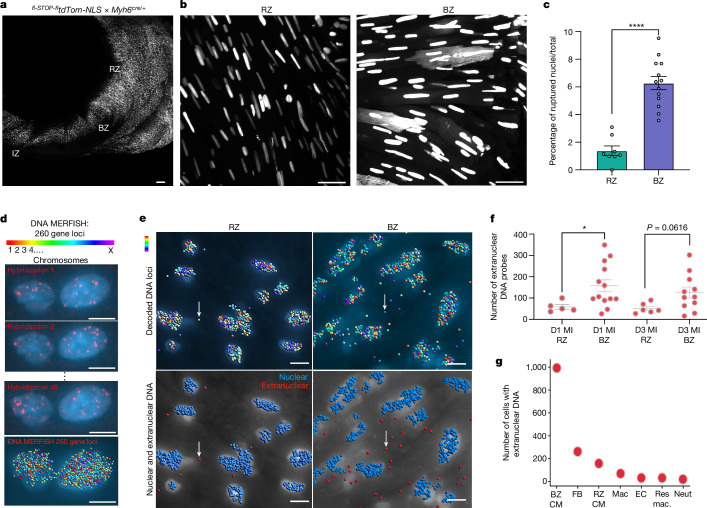
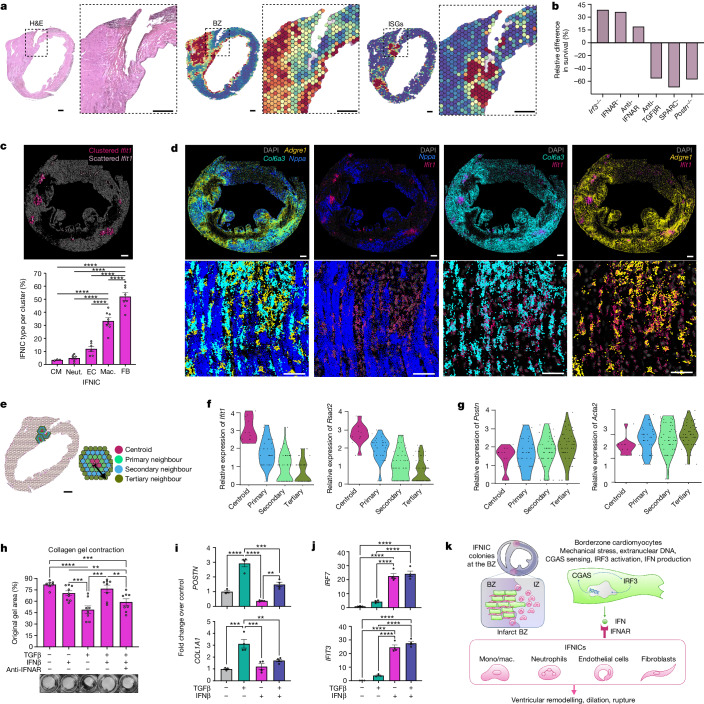
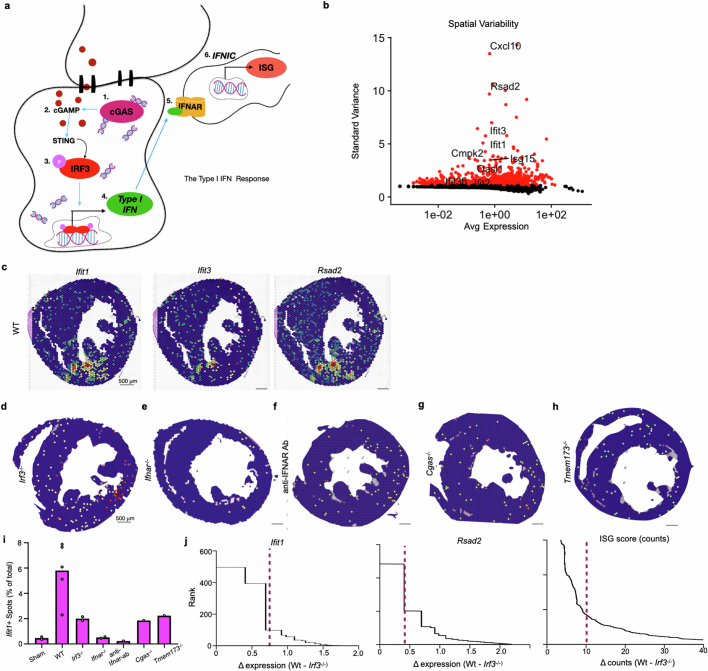
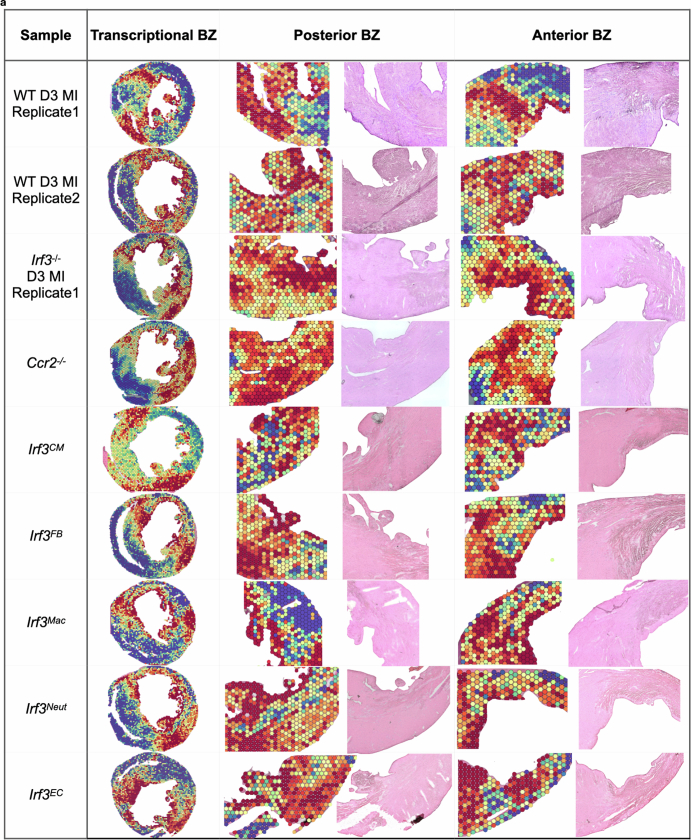
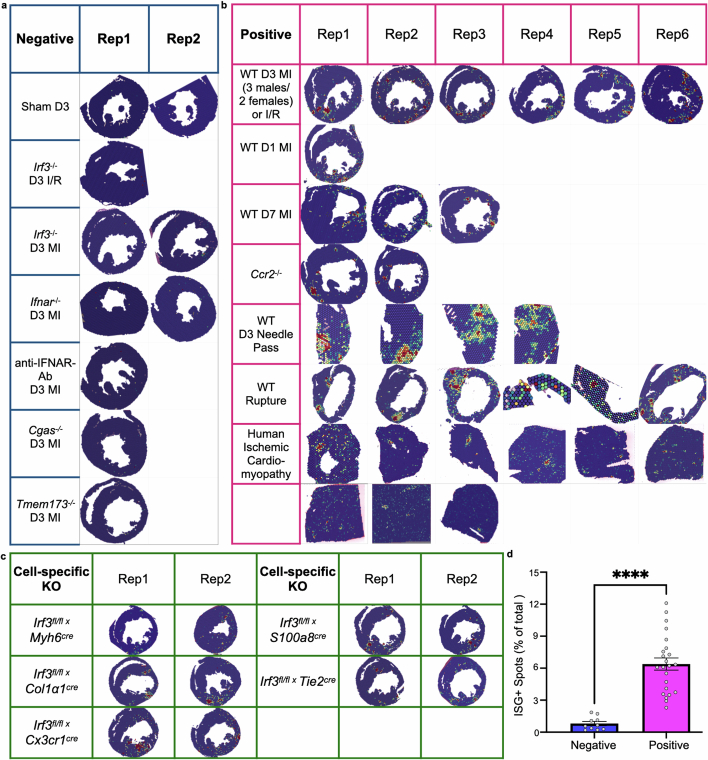
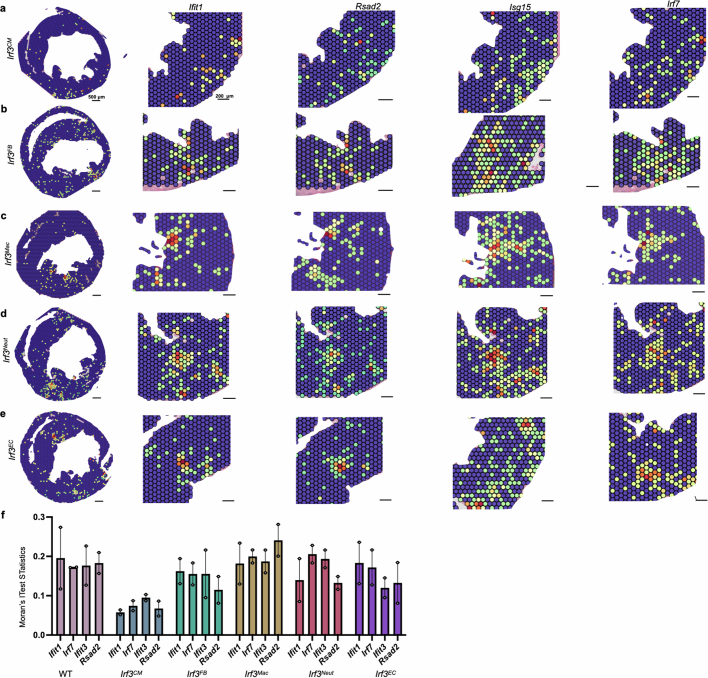
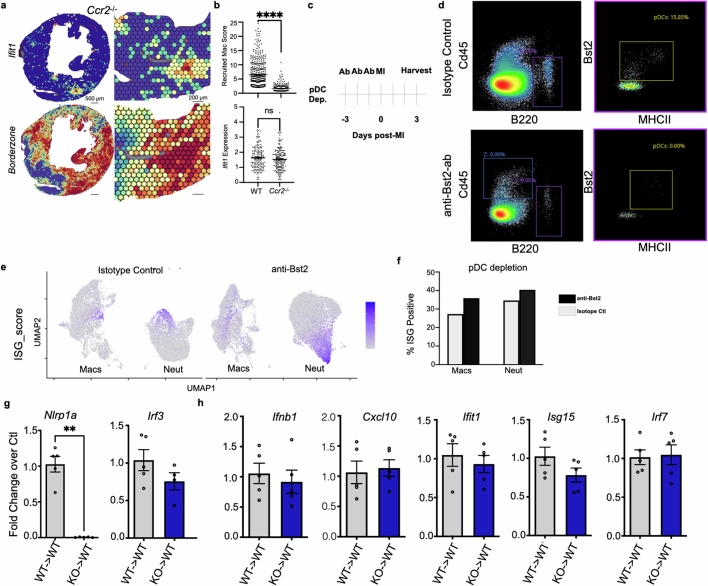
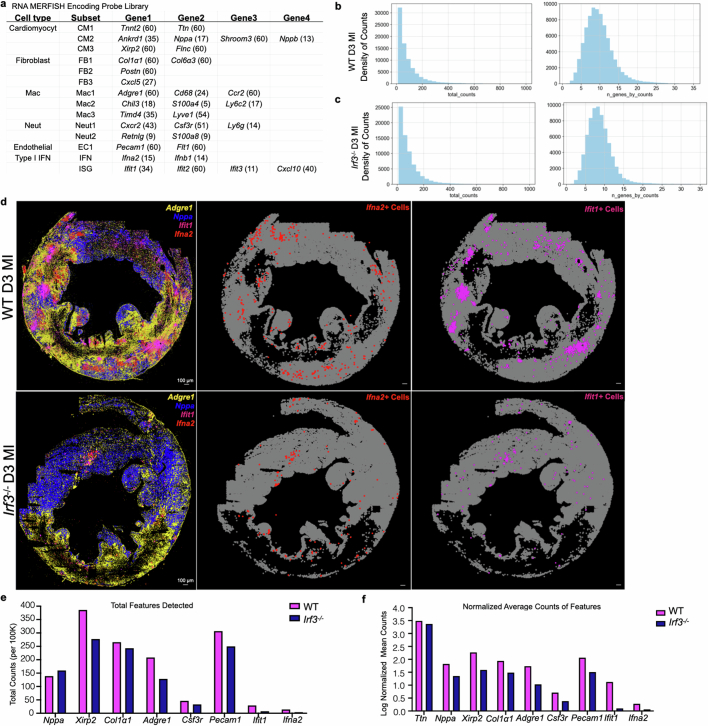
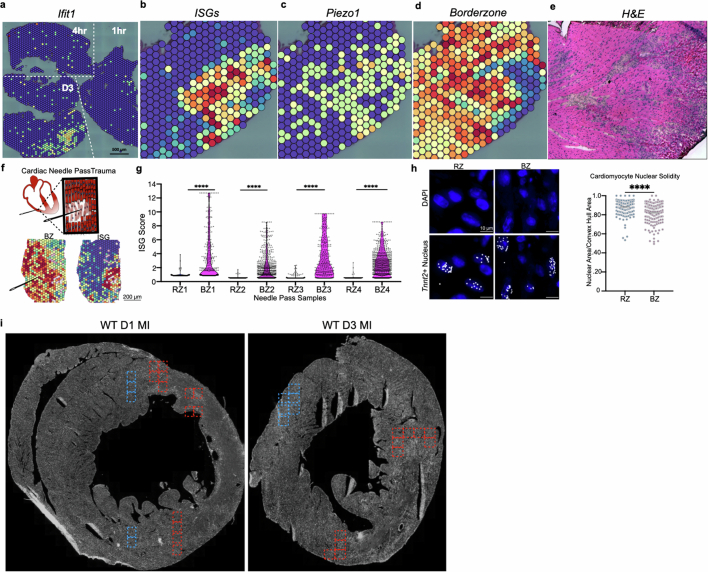
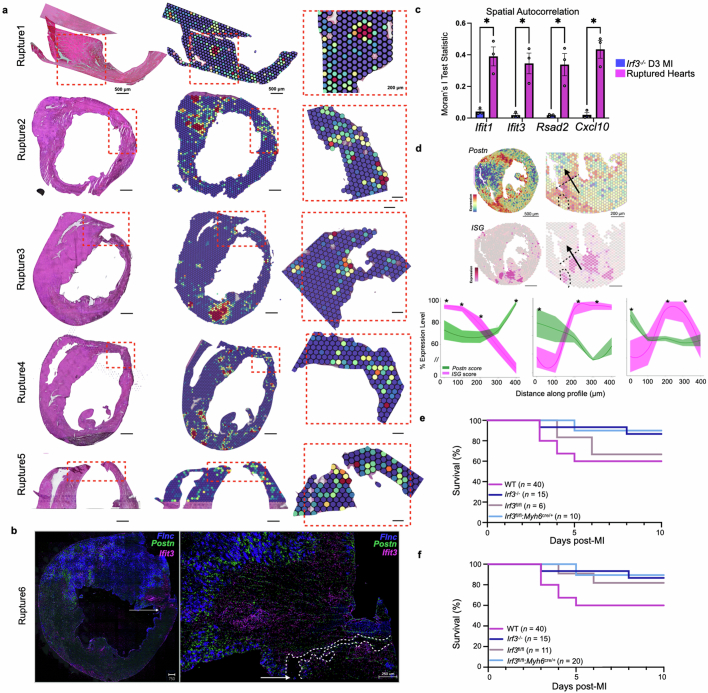
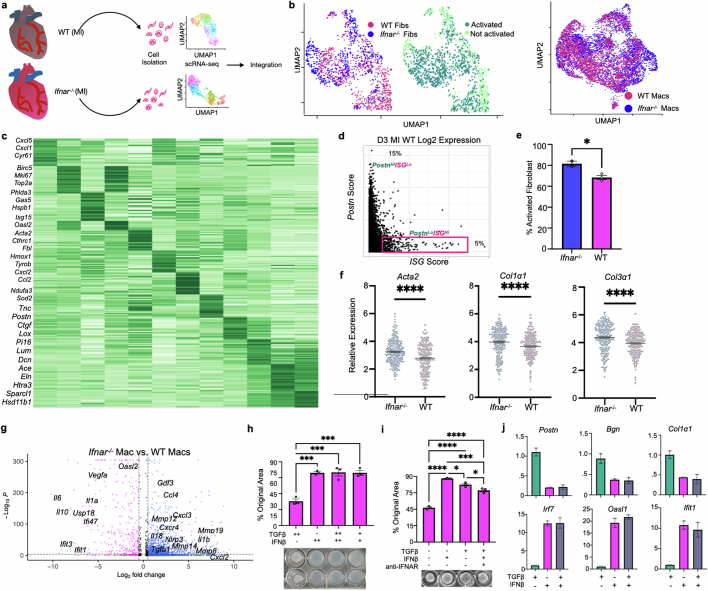
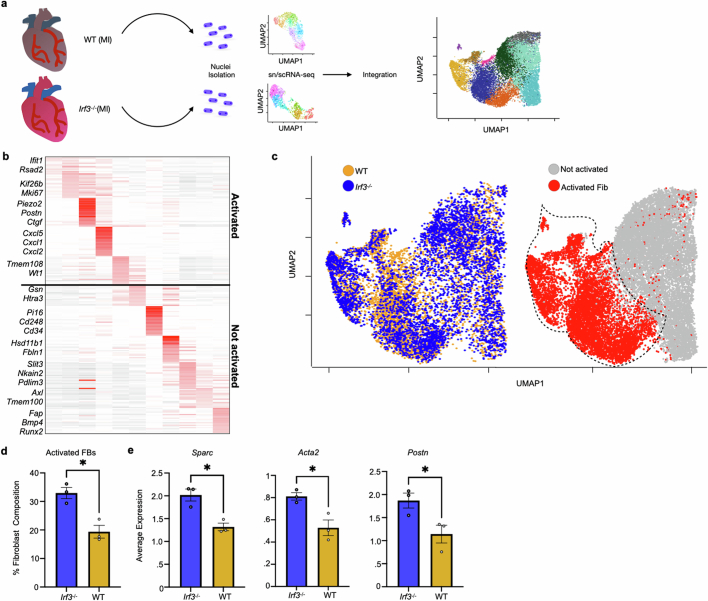
Sterile inflammation after myocardial infarction is classically credited to myeloid cells interacting with dead cell debris in the infarct zone1,2. Here we show that cardiomyocytes are the dominant initiators of a previously undescribed type I interferon response in the infarct borderzone. Using spatial transcriptomics analysis in mice and humans, we find that myocardial infarction induces colonies of interferon-induced cells (IFNICs) expressing interferon-stimulated genes decorating the borderzone, where cardiomyocytes experience mechanical stress, nuclear rupture and escape of chromosomal DNA. Cardiomyocyte-selective deletion of Irf3 abrogated IFNIC colonies, whereas mice lacking Irf3 in fibroblasts, macrophages, neutrophils or endothelial cells, Ccr2-deficient mice or plasmacytoid-dendritic-cell-depleted mice did not. Interferons blunted the protective matricellular programs and contractile function of borderzone fibroblasts, and increased vulnerability to pathological remodelling. In mice that died after myocardial infarction, IFNIC colonies were immediately adjacent to sites of ventricular rupture, while mice lacking IFNICs were protected from rupture and exhibited improved survival3. Together, these results reveal a pathological borderzone niche characterized by a cardiomyocyte-initiated innate immune response. We suggest that selective inhibition of IRF3 activation in non-immune cells could limit ischaemic cardiomyopathy while avoiding broad immunosuppression.
© 2024. The Author(s).
Conflict of interest statement
K.L.C. is a cofounder, board member and consultant for, and holds equity interest in Ventrix and Karios Technologies. K.R.K. and V.K.N. hold a patent declaration PCT 114198-5210 on compositions and methods to treat muscle injury and disease.
Figures
Comment in
-
Interferon response at the border zone of the infarcted heart.Nat Rev Cardiol. 2024 Nov;21(11):740. doi: 10.1038/s41569-024-01078-y. Nat Rev Cardiol. 2024. PMID: 39289539 No abstract available.
