

Fate induction in CD8 CAR T cells through asymmetric cell division
- PMID: 39198645
- PMCID: PMC11410665
- DOI: 10.1038/s41586-024-07862-7
Fate induction in CD8 CAR T cells through asymmetric cell division
Abstract
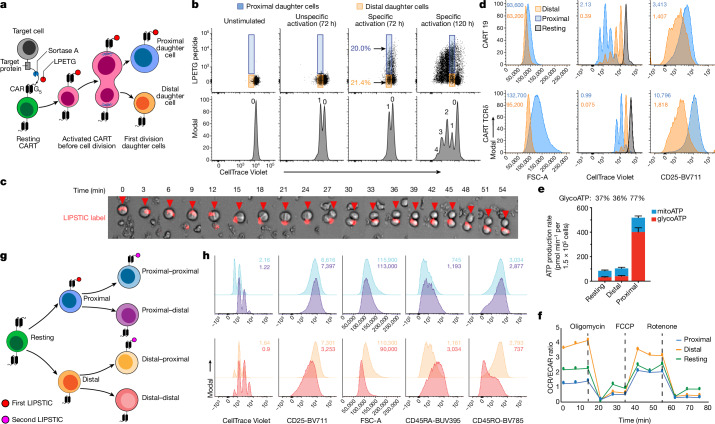
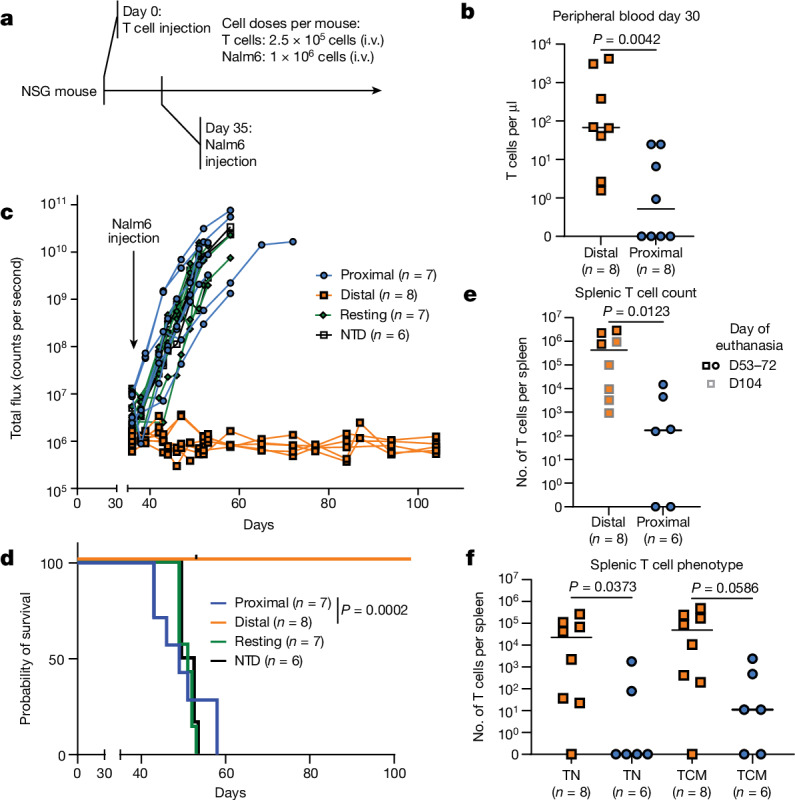
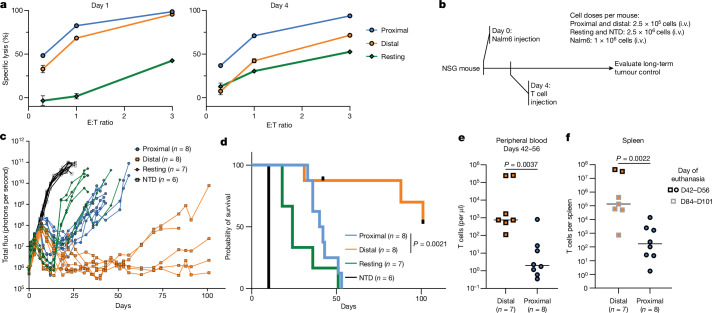
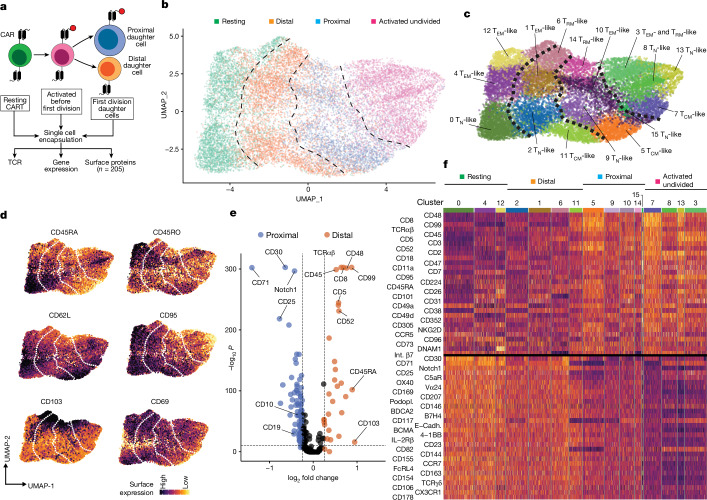
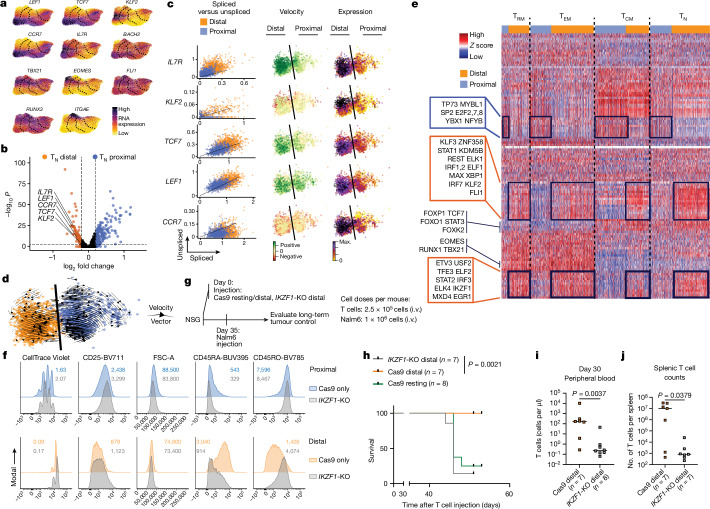
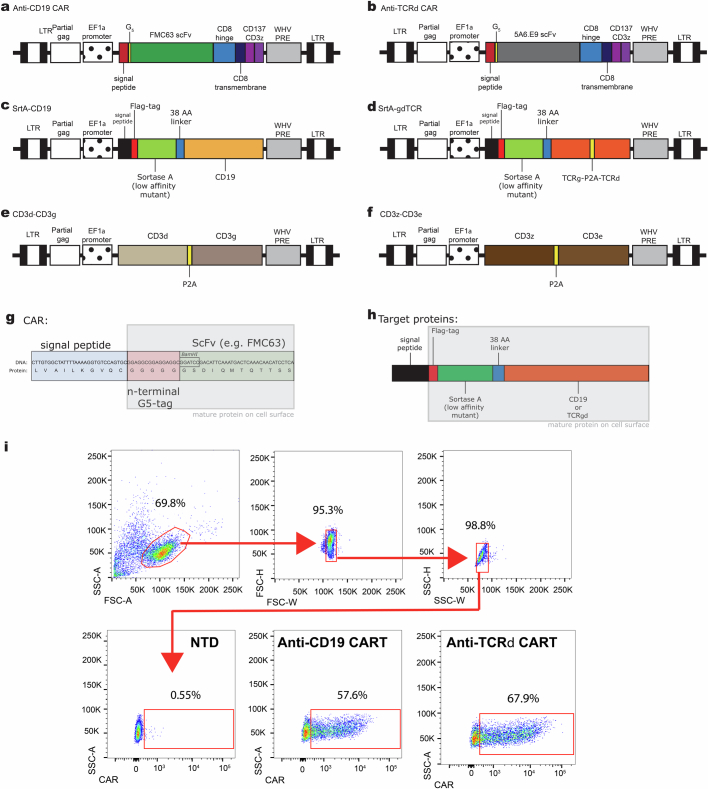
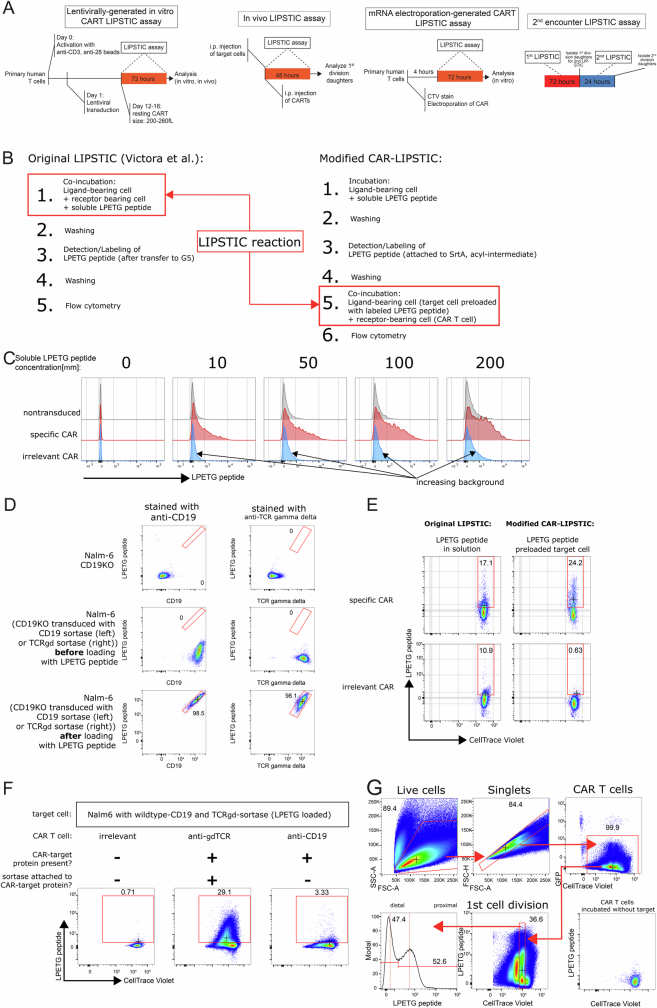
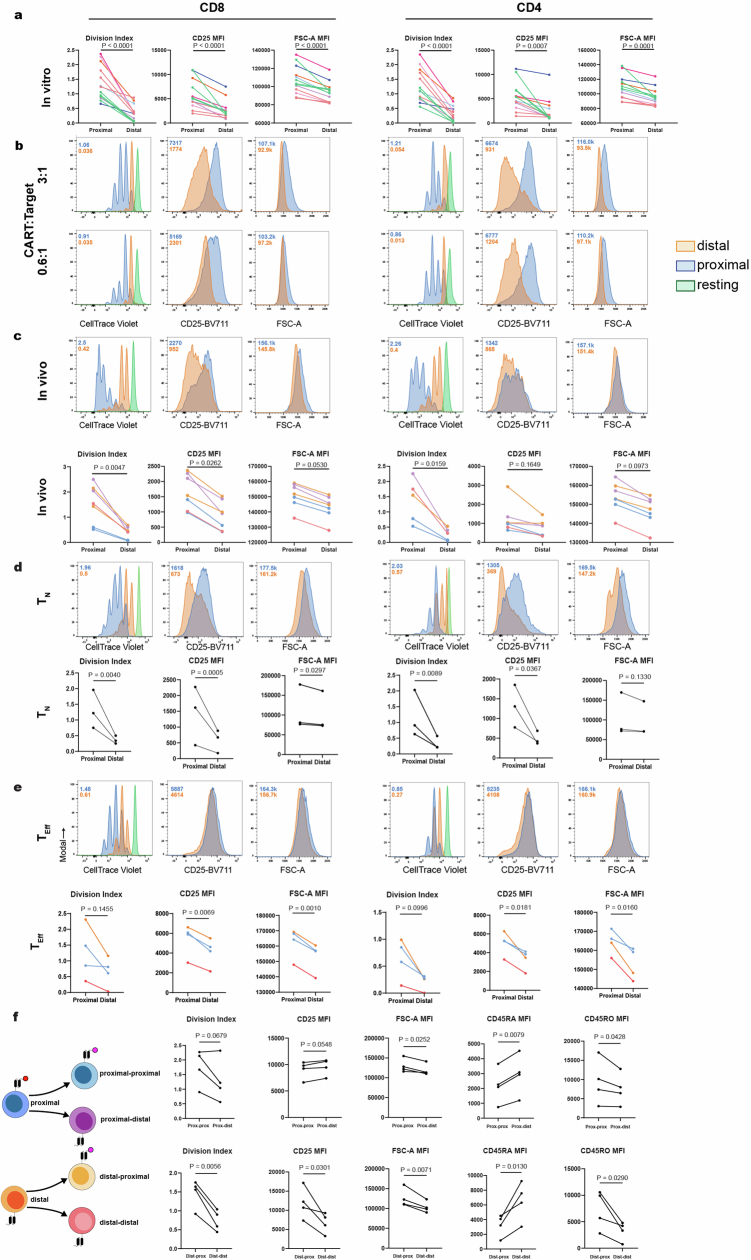
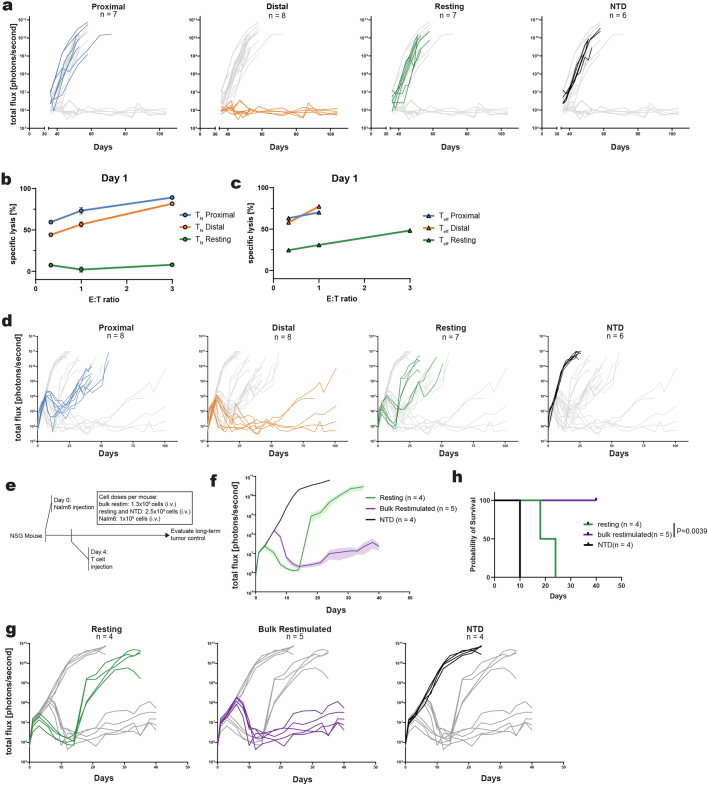
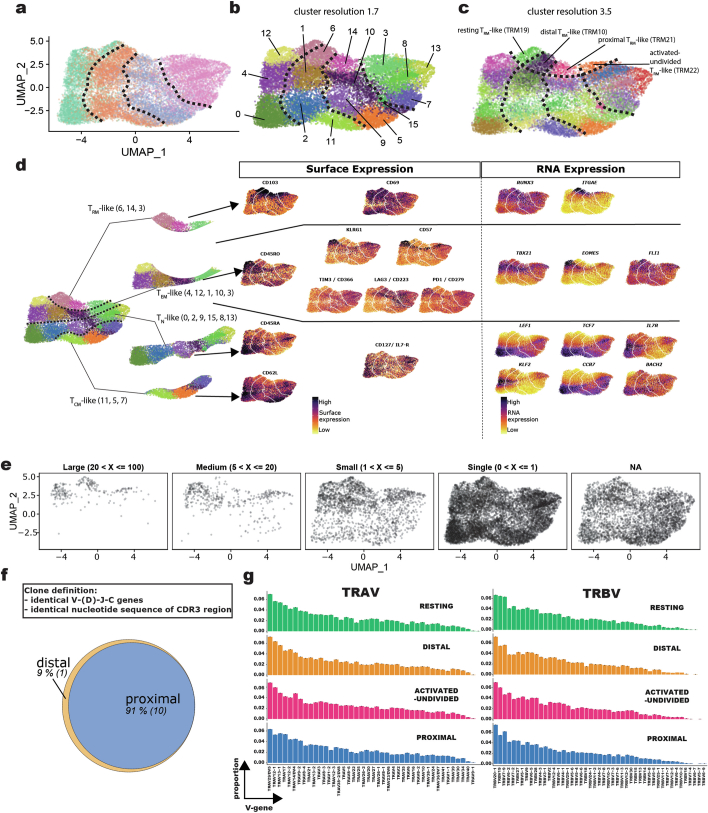
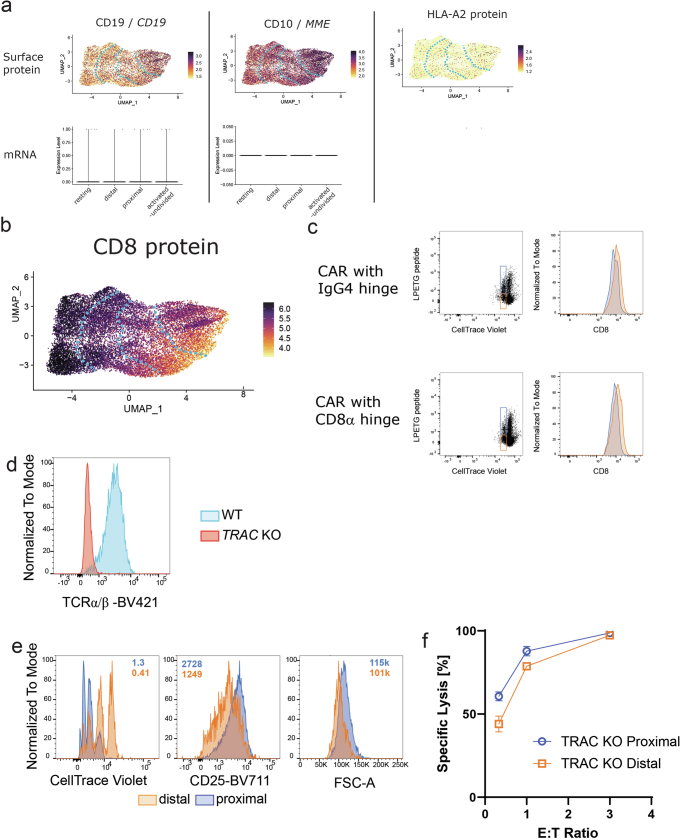
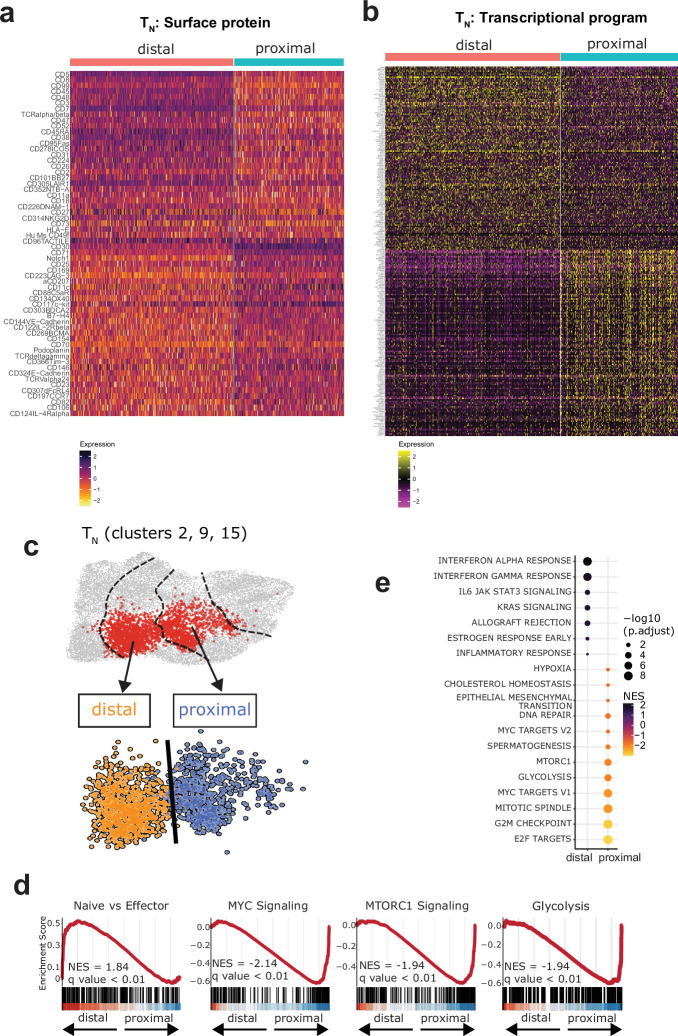
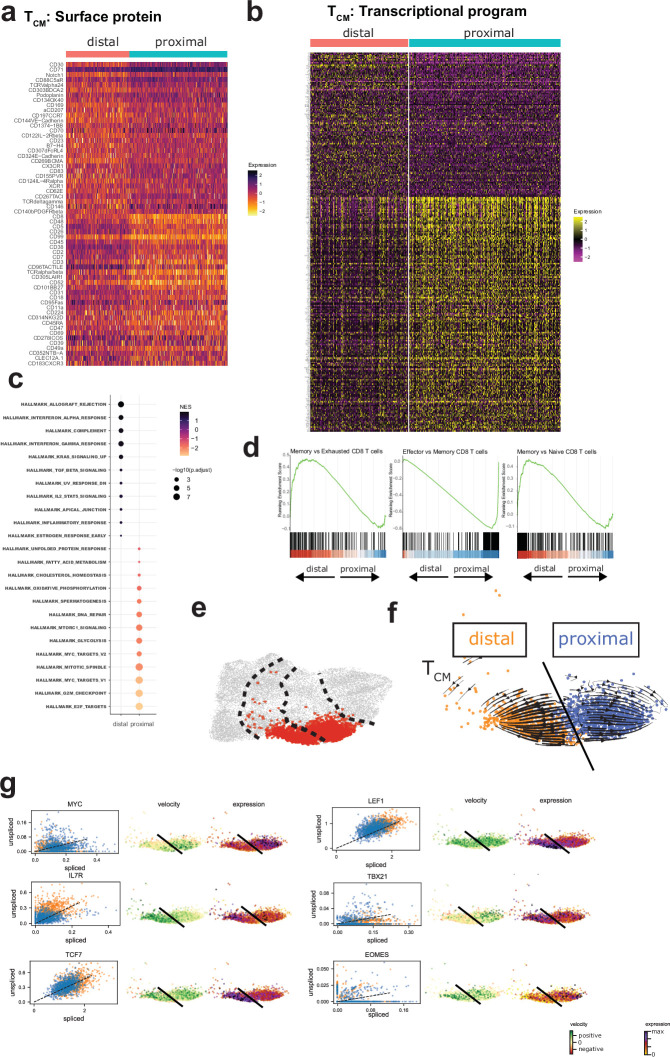
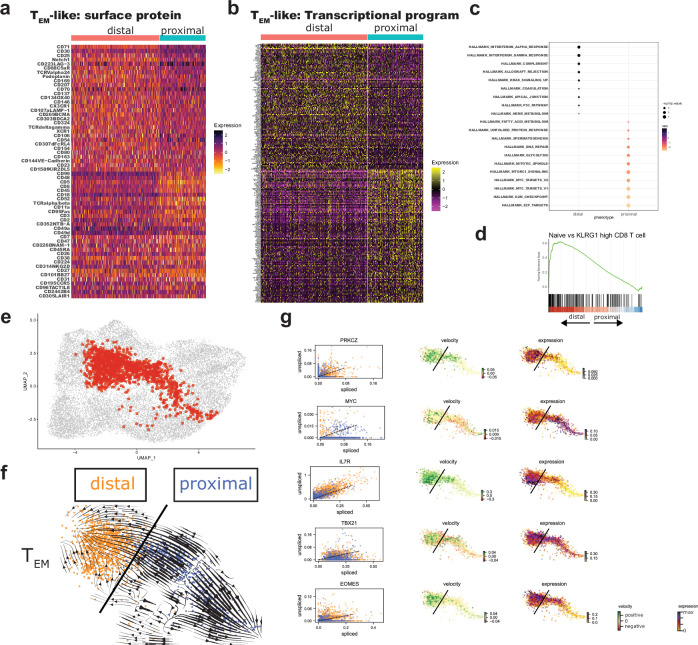
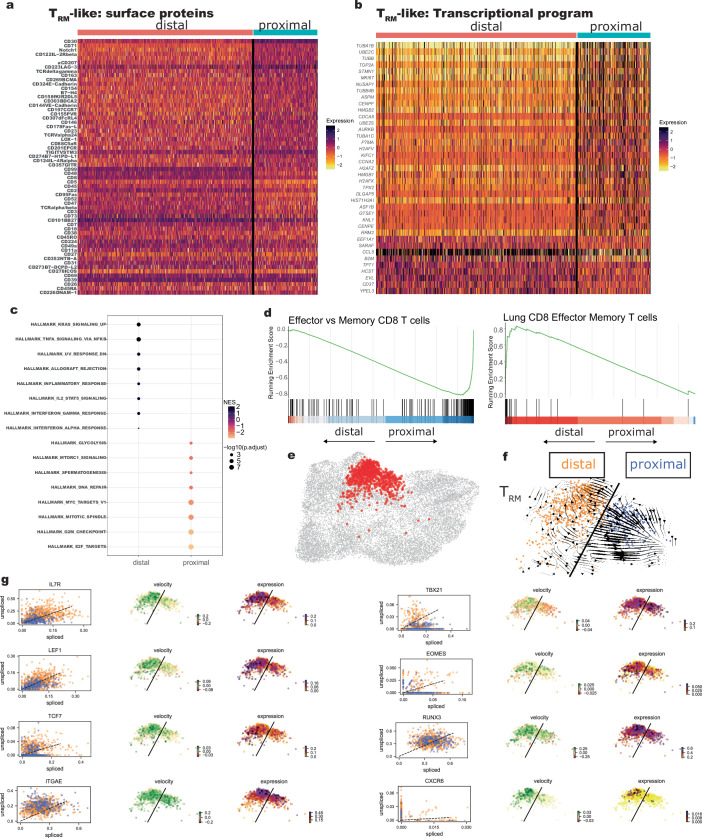
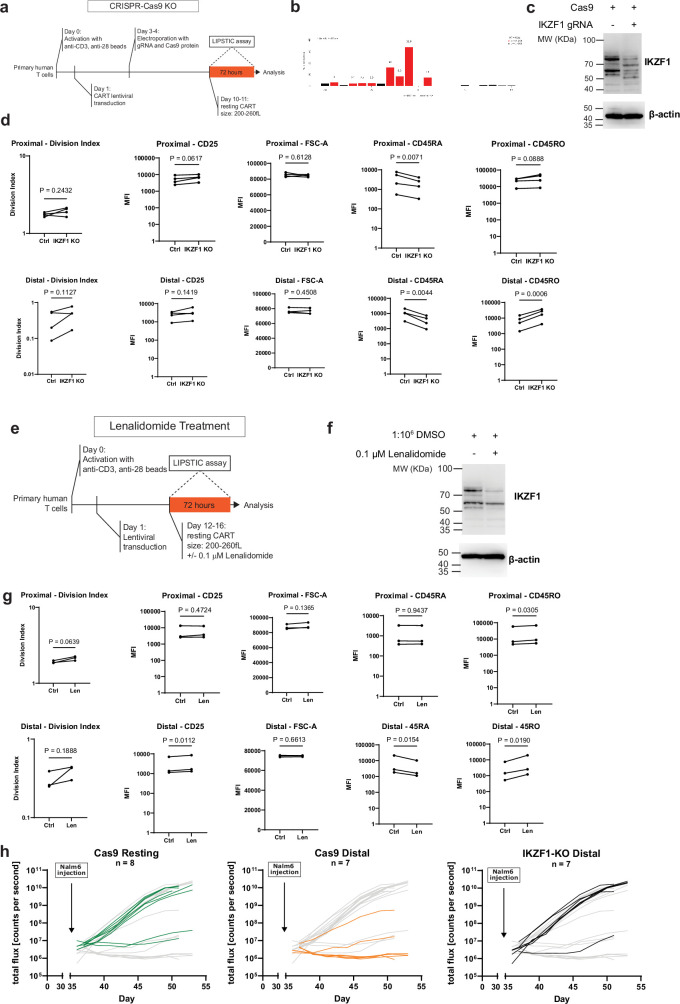
Early expansion and long-term persistence predict efficacy of chimeric antigen receptor T cells (CARTs)1-7, but mechanisms governing effector versus memory CART differentiation and whether asymmetric cell division induces differential fates in human CARTs remain unclear. Here we show that target-induced proximity labelling enables isolation of first-division proximal-daughter and distal-daughter CD8 CARTs that asymmetrically distribute their surface proteome and transcriptome, resulting in divergent fates. Target-engaged CARs remain on proximal daughters, which inherit a surface proteome resembling activated-undivided CARTs, whereas the endogenous T cell receptor and CD8 enrich on distal daughters, whose surface proteome resembles resting CARTs, correlating with glycolytic and oxidative metabolism, respectively. Despite memory-precursor phenotype and in vivo longevity, distal daughters demonstrate transient potent cytolytic activity similar to proximal daughters, uncovering an effector-like state in distal daughters destined to become memory CARTs. Both partitioning of pre-existing transcripts and changes in RNA velocity contribute to asymmetry of fate-determining factors, resulting in diametrically opposed transcriptional trajectories. Independent of naive, memory or effector surface immunophenotype, proximal-daughter CARTs use core sets of transcription factors known to support proliferation and effector function. Conversely, transcription factors enriched in distal daughters restrain differentiation and promote longevity, evidenced by diminished long-term in vivo persistence and function of distal-daughter CARTs after IKZF1 disruption. These studies establish asymmetric cell division as a framework for understanding mechanisms of CART differentiation and improving therapeutic outcomes.
© 2024. The Author(s).
Conflict of interest statement
C.T.E.—equity (Cabaletta Bio), patent licensing (Cabaletta Bio, Novartis). A.S.P.—equity, payments, research grants (Cabaletta Bio), patent licensing (Cabaletta Bio, Novartis), consultant (Janssen). S.O.—patent licensing (Cabaletta Bio). R.O.C.—equity (Nucleus Biologics), research grant (NeoTx) and patent licensing (Novartis). The remaining authors declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
