

BDNF-Regulated Modulation of Striatal Circuits and Implications for Parkinson's Disease and Dystonia
- PMID: 39200225
- PMCID: PMC11351984
- DOI: 10.3390/biomedicines12081761
BDNF-Regulated Modulation of Striatal Circuits and Implications for Parkinson's Disease and Dystonia
Abstract
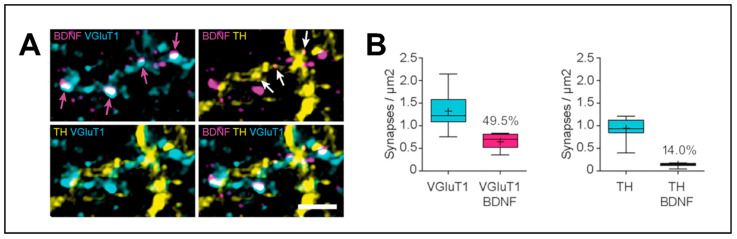
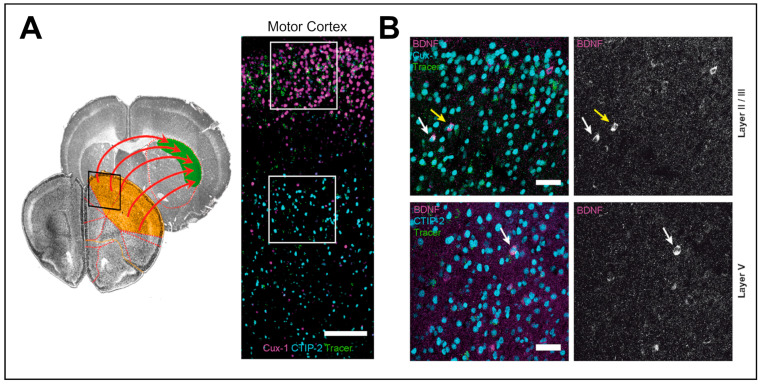
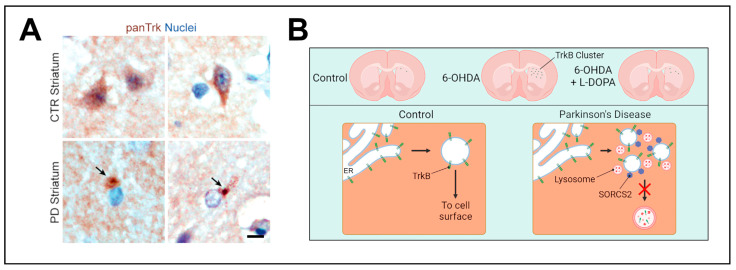
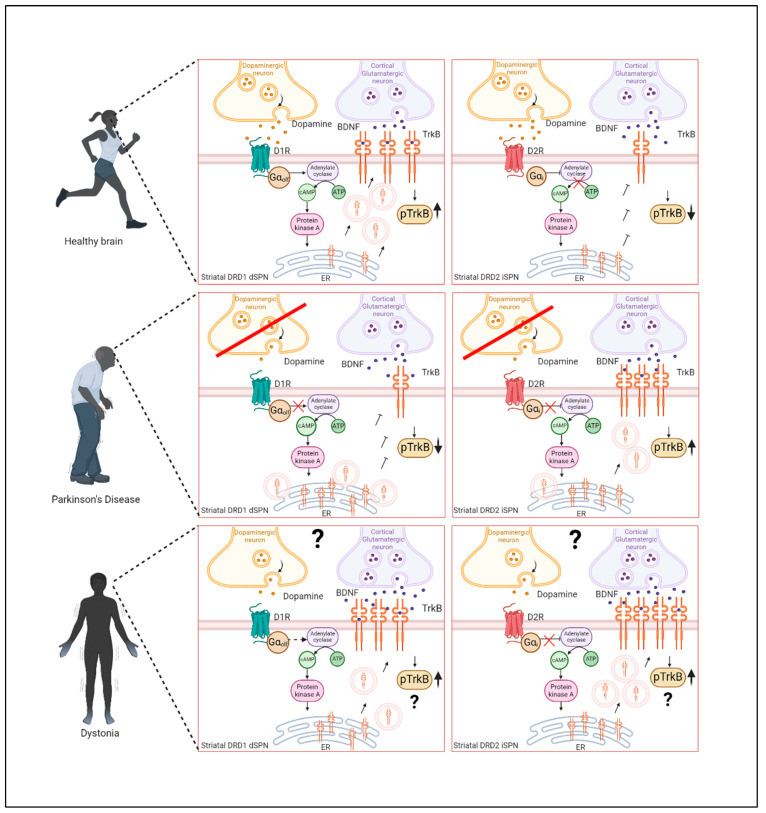
Neurotrophins, particularly brain-derived neurotrophic factor (BDNF), act as key regulators of neuronal development, survival, and plasticity. BDNF is necessary for neuronal and functional maintenance in the striatum and the substantia nigra, both structures involved in the pathogenesis of Parkinson's Disease (PD). Depletion of BDNF leads to striatal degeneration and defects in the dendritic arborization of striatal neurons. Activation of tropomyosin receptor kinase B (TrkB) by BDNF is necessary for the induction of long-term potentiation (LTP), a form of synaptic plasticity, in the hippocampus and striatum. PD is characterized by the degeneration of nigrostriatal neurons and altered striatal plasticity has been implicated in the pathophysiology of PD motor symptoms, leading to imbalances in the basal ganglia motor pathways. Given its essential role in promoting neuronal survival and meditating synaptic plasticity in the motor system, BDNF might have an important impact on the pathophysiology of neurodegenerative diseases, such as PD. In this review, we focus on the role of BDNF in corticostriatal plasticity in movement disorders, including PD and dystonia. We discuss the mechanisms of how dopaminergic input modulates BDNF/TrkB signaling at corticostriatal synapses and the involvement of these mechanisms in neuronal function and synaptic plasticity. Evidence for alterations of BDNF and TrkB in PD patients and animal models are reviewed, and the potential of BDNF to act as a therapeutic agent is highlighted. Advancing our understanding of these mechanisms could pave the way toward innovative therapeutic strategies aiming at restoring neuroplasticity and enhancing motor function in these diseases.
Keywords: BDNF; PD; Parkinson’s disease; TrkB; brain-derived neurotrophic factor; dopamine; dystonia; neurological diseases; neuronal plasticity.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
References
Publication types
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
