

Computational Analyses Reveal Deregulated Clock Genes Associated with Breast Cancer Development in Night Shift Workers
- PMID: 39201344
- PMCID: PMC11355052
- DOI: 10.3390/ijms25168659
Computational Analyses Reveal Deregulated Clock Genes Associated with Breast Cancer Development in Night Shift Workers
Abstract
Breast cancer (BC) is the leading cause of cancer death among women worldwide. Women employed in shift jobs face heightened BC risk due to prolonged exposure to night shift work (NSW), classified as potentially carcinogenic by the International Agency for Research on Cancer (IARC). This risk is linked to disruptions in circadian rhythms governed by clock genes at the cellular level. However, the molecular mechanisms are unclear. This study aimed to assess clock genes as potential BC biomarkers among women exposed to long-term NSW. Clock gene expression was analysed in paired BC and normal breast tissues within Nurses' Health Studies I and II GEO datasets. Validation was performed on additional gene expression datasets from healthy night shift workers and women with varying BC susceptibility, as well as single-cell sequencing datasets. Post-transcriptional regulators of clock genes were identified through miRNA analyses. Significant alterations in clock gene expression in BC compared to normal tissues were found. BHLHE40, CIART, CLOCK, PDPK1, and TIMELESS were over-expressed, while HLF, NFIL3, NPAS3, PER1, PER3, SIM1, and TEF were under-expressed. The downregulation of PER1 and TEF and upregulation of CLOCK correlated with increased BC risk in healthy women. Also, twenty-six miRNAs, including miR-10a, miR-21, miR-107, and miR-34, were identified as potential post-transcriptional regulators influenced by NSW. In conclusion, a panel of clock genes and circadian miRNAs are suggested as BC susceptibility biomarkers among night shift workers, supporting implications for risk stratification and early detection strategies.
Keywords: biomarkers; breast cancer; circadian rhythms; clock genes; computational study; micro-RNAs; night shift work.
Conflict of interest statement
The authors declare no conflicts of interest.
Figures
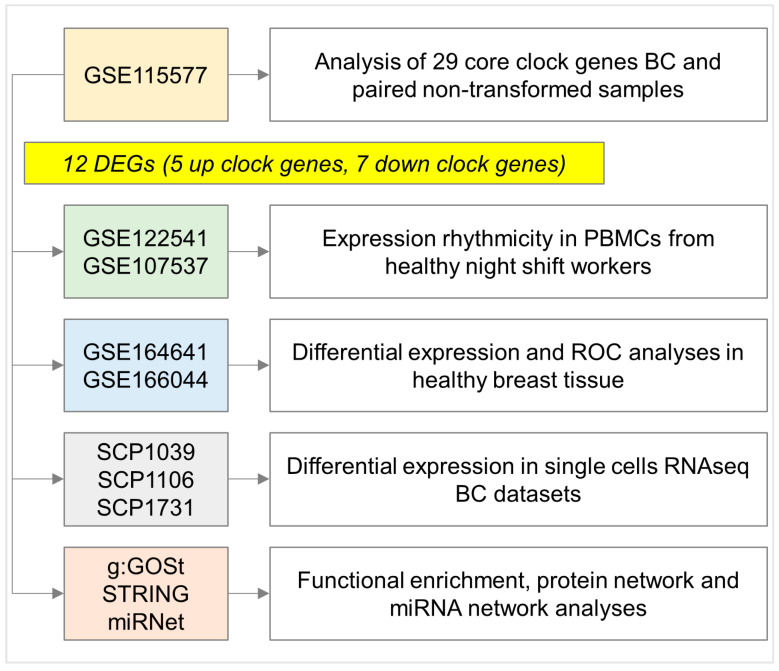
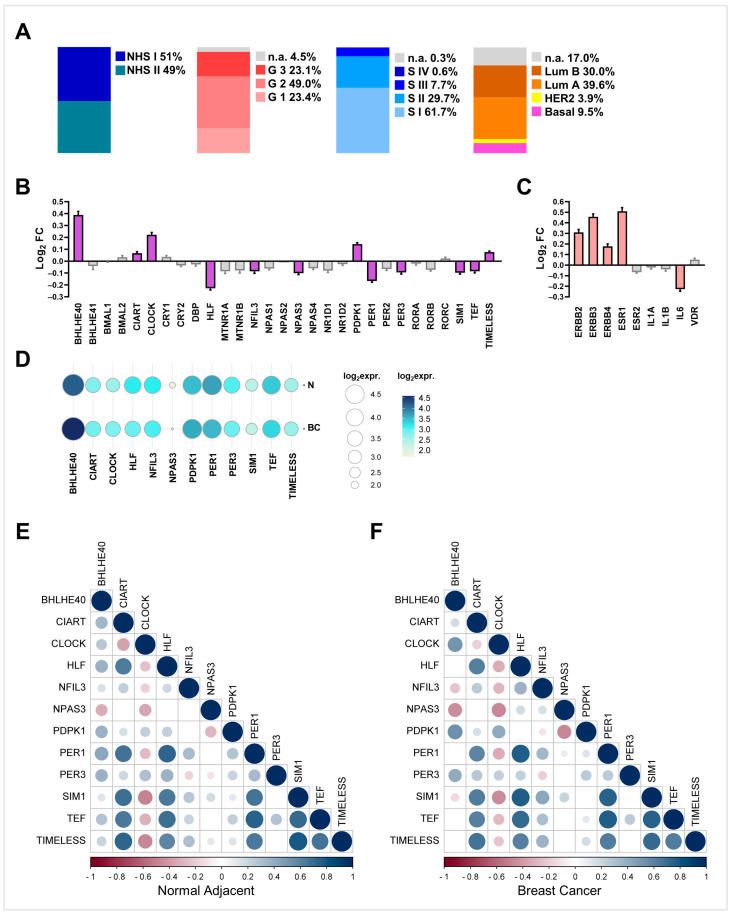
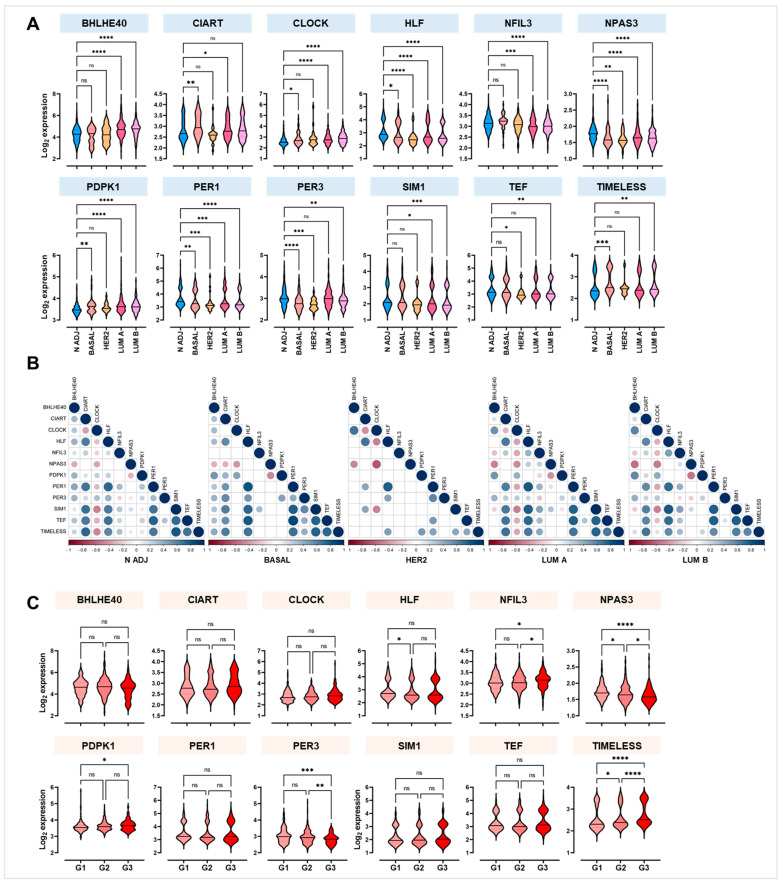
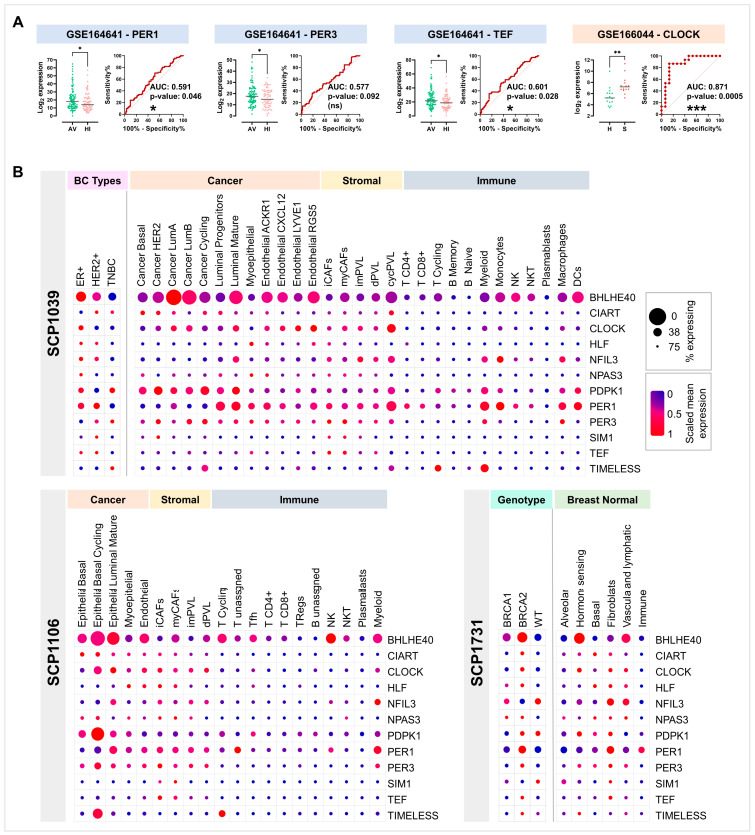
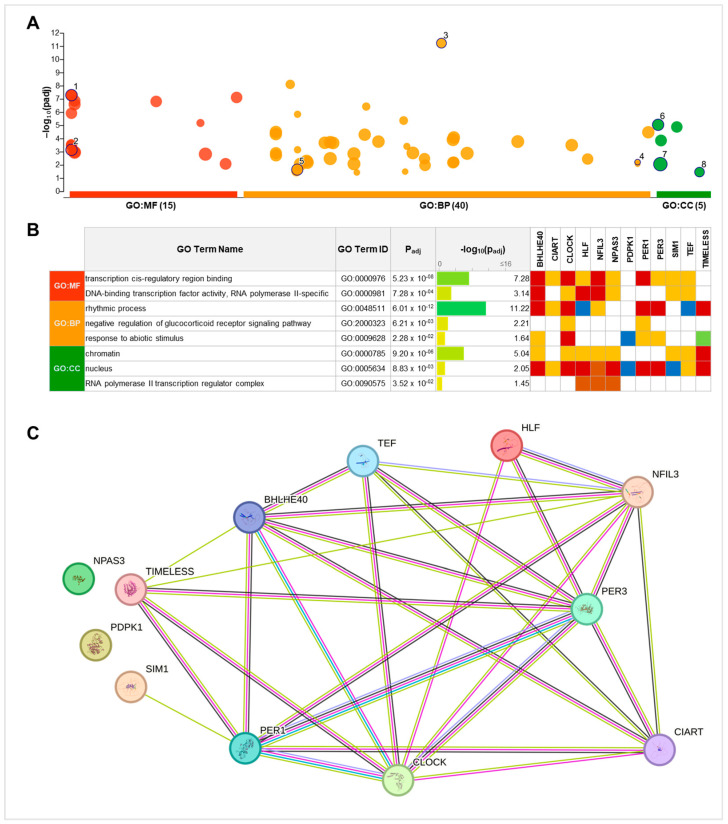
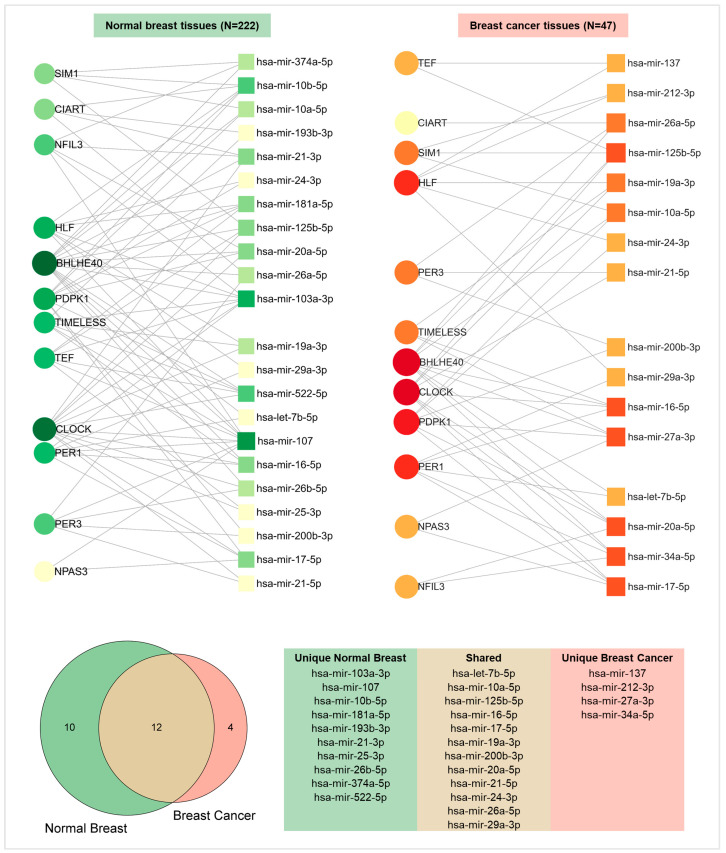
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
