

Human Stimulator of Interferon Genes Promotes Rhinovirus C Replication in Mouse Cells In Vitro and In Vivo
- PMID: 39205256
- PMCID: PMC11358906
- DOI: 10.3390/v16081282
Human Stimulator of Interferon Genes Promotes Rhinovirus C Replication in Mouse Cells In Vitro and In Vivo
Abstract
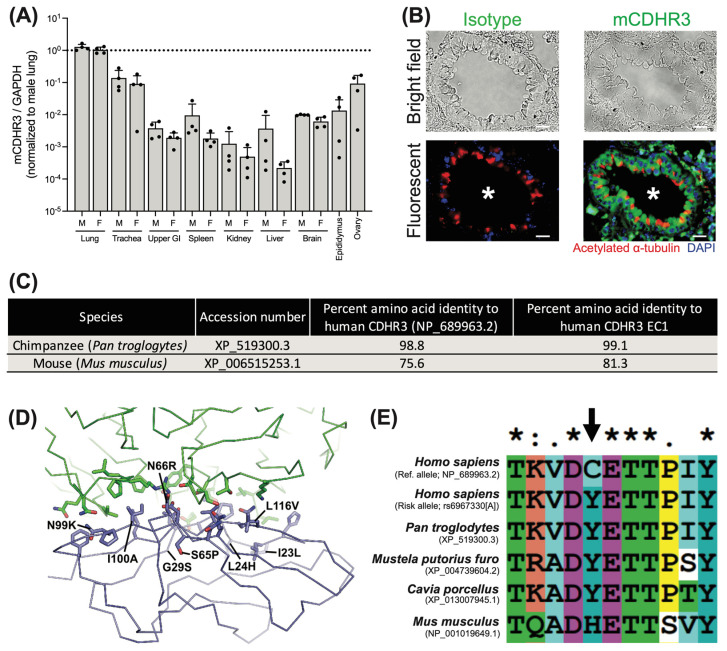
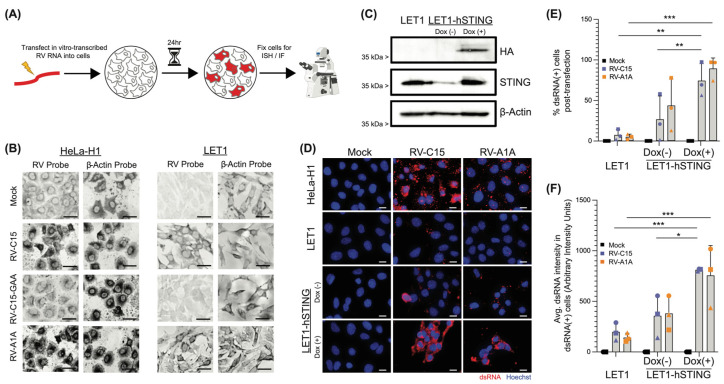
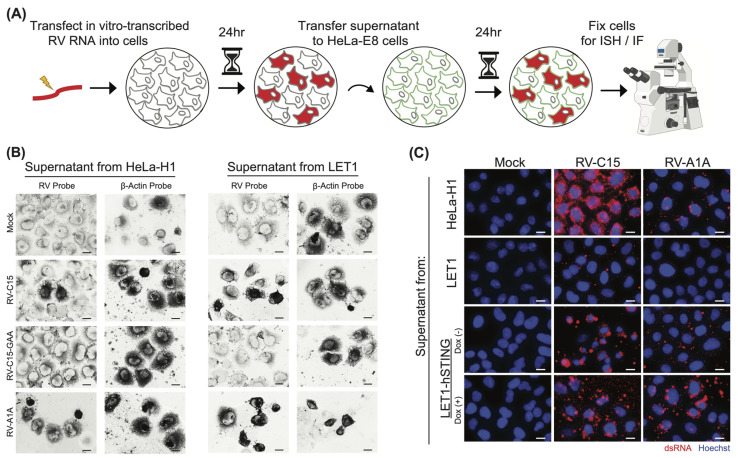
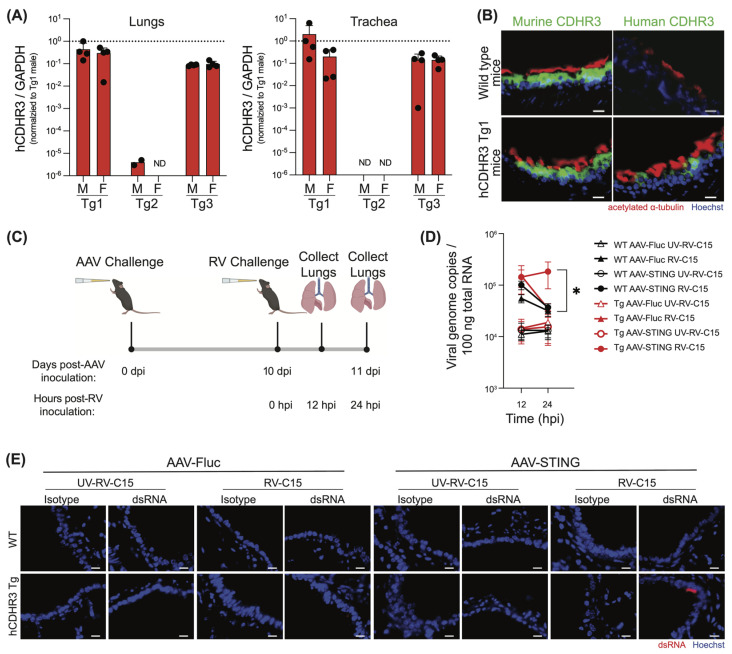
Rhinovirus C (RV-C) infects airway epithelial cells and is an important cause of acute respiratory disease in humans. To interrogate the mechanisms of RV-C-mediated disease, animal models are essential. Towards this, RV-C infection was recently reported in wild-type (WT) mice, yet, titers were not sustained. Therefore, the requirements for RV-C infection in mice remain unclear. Notably, prior work has implicated human cadherin-related family member 3 (CDHR3) and stimulator of interferon genes (STING) as essential host factors for virus uptake and replication, respectively. Here, we report that even though human (h) and murine (m) CDHR3 orthologs have similar tissue distribution, amino acid sequence homology is limited. Further, while RV-C can replicate in mouse lung epithelial type 1 (LET1) cells and produce infectious virus, we observed a significant increase in the frequency and intensity of dsRNA-positive cells following hSTING expression. Based on these findings, we sought to assess the impact of hCDHR3 and hSTING on RV-C infection in mice in vivo. Thus, we developed hCDHR3 transgenic mice, and utilized adeno-associated virus (AAV) to deliver hSTING to the murine airways. Subsequent challenge of these mice with RV-C15 revealed significantly higher titers 24 h post-infection in mice expressing both hCDHR3 and hSTING-compared to either WT mice, or mice with hCDHR3 or hSTING alone, indicating more efficient infection. Ultimately, this mouse model can be further engineered to establish a robust in vivo model, recapitulating viral dynamics and disease.
Keywords: CDHR3; STING; mouse model; rhinovirus C.
Conflict of interest statement
M.A.S. received funding from AstraZeneca PLC to support the studies reported here.
Figures
References
-
- Matsuno A.K., Gagliardi T.B., Paula F.E., Luna L.K.S., Jesus B.L.S., Stein R.T., Aragon D.C., Carlotti A.P.C.P., Arruda E. Human coronavirus alone or in co-infection with rhinovirus C is a risk factor for severe respiratory disease and admission to the pediatric intensive care unit: A one-year study in Southeast Brazil. PLoS ONE. 2019;14:e0217744. doi: 10.1371/journal.pone.0217744. - DOI - PMC - PubMed
-
- Ng K.T., Oong X.Y., Lim S.H., Chook J.B., Takebe Y., Chan Y.F., Chan K.G., Hanafi N.S., Pang Y.K., Kamarulzaman A., et al. Viral Load and Sequence Analysis Reveal the Symptom Severity, Diversity, and Transmission Clusters of Rhinovirus Infections. Clin. Infect. Dis. 2018;67:261–268. doi: 10.1093/cid/ciy063. - DOI - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
