

C9orf72 polyPR interaction with the nuclear pore complex
- PMID: 39205388
- PMCID: PMC11495645
- DOI: 10.1016/j.bpj.2024.08.024
C9orf72 polyPR interaction with the nuclear pore complex
Abstract
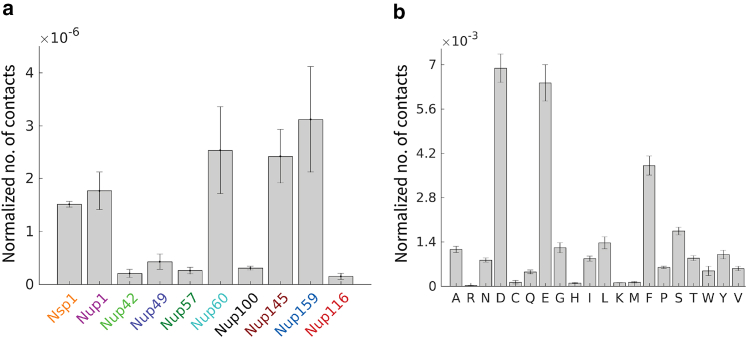
The C9orf72 gene associated with amyotrophic lateral sclerosis/frontotemporal dementia is translated to five dipeptide repeat proteins, among which poly-proline-arginine (PR) is the most toxic in cell and animal models, contributing to a variety of cellular defects. It has been proposed that polyPR disrupts nucleocytoplasmic transport (NCT) through several mechanisms including accumulation in the nuclear pore complex (NPC), accumulation in the nucleolus, and direct interactions with transport receptors. The NPC, which is the key regulator of transport between the cytoplasm and nucleus, plays a central role in these suggested mechanisms. Exploring polyPR interaction with the NPC provides valuable insight into the molecular details of polyPR-mediated NCT defects. To address this, we use coarse-grained molecular dynamics models of polyPR and the yeast NPC lined with intrinsically disordered FG-nucleoporins (FG-Nups). Our findings indicate no aggregation of polyPR within the NPC or permanent binding to FG-Nups. Instead, polyPR translocates through the NPC, following a trajectory through the central low-density region of the pore. In the case of longer polyPRs, we observe a higher energy barrier for translocation and a narrower translocation channel. Our study shows that polyPR and FG-Nups are mainly engaged in steric interactions inside the NPC with only a small contribution of specific cation-pi, hydrophobic, and electrostatic interactions, allowing polyPR to overcome the entropic barrier of the NPC in a size-dependent manner.
Copyright © 2024 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors do not declare any conflicts of interest.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
