

Thrombomodulin (p.Cys537Stop) is released from cells by an unusual membrane insertion/leakage mechanism
- PMID: 39208365
- PMCID: PMC11532747
- DOI: 10.1182/bloodadvances.2024013546
Thrombomodulin (p.Cys537Stop) is released from cells by an unusual membrane insertion/leakage mechanism
Abstract
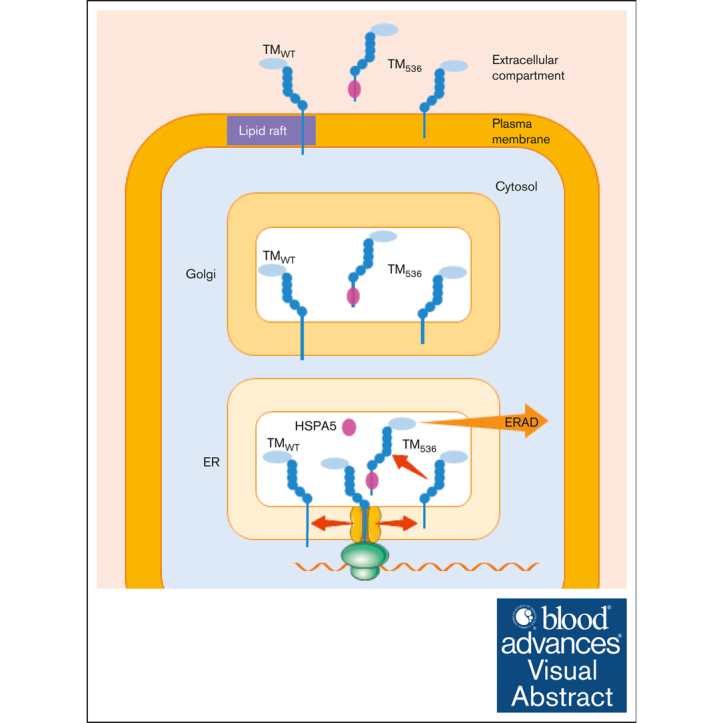
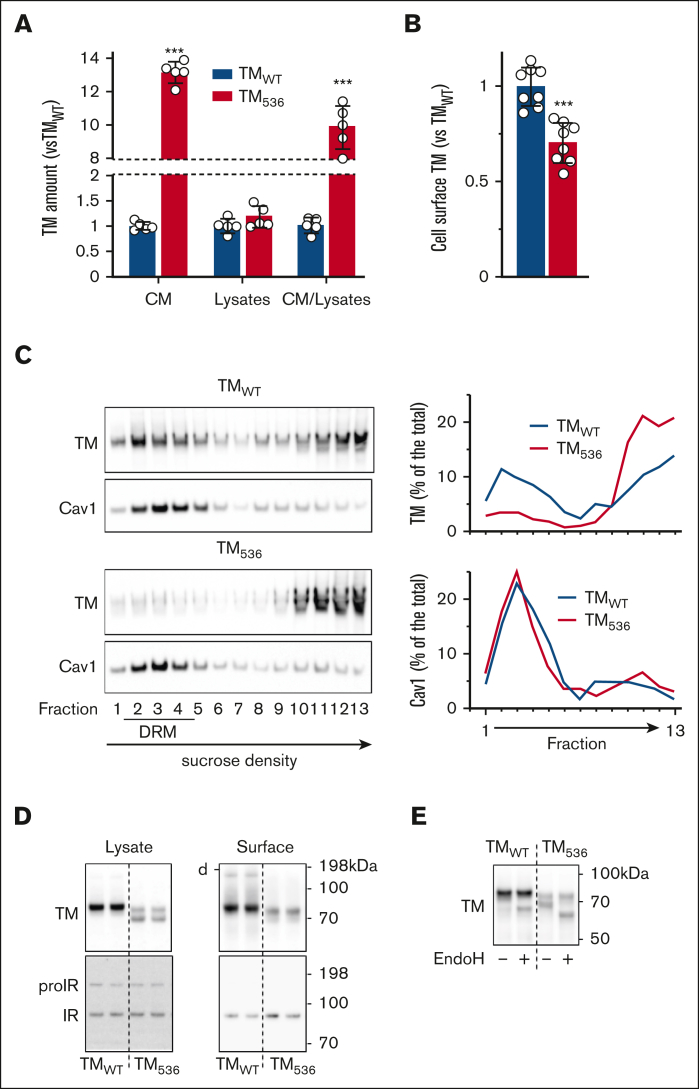
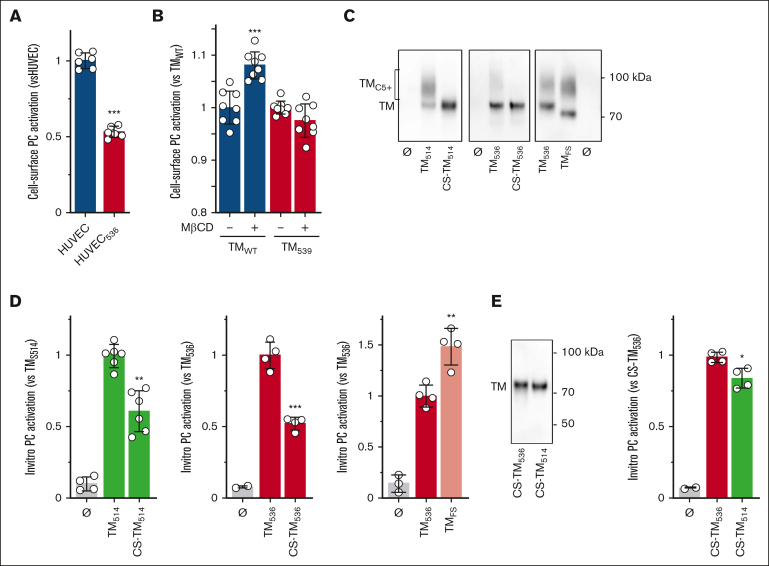
Expression of the thrombomodulin (TM) variant c.1611C>A (p.Cys537Stop) leads to the synthesis of a protein with no cytoplasmic tail and a transmembrane domain shortened by 3 amino acids (TM536). However, little is known regarding the release mechanism and properties of TM536. Using umbilical vein endothelial cells and peripheral blood-derived endothelial colony-forming cells from a heterozygous carrier of the TM536 variant as well as overexpression cell models, we demonstrated that TM536 is released from cells by an unusual mechanism. First, TM536 is inserted into the endoplasmic reticulum (ER) membrane, then, because of the low hydrophobicity of its intramembrane domain, it escapes from it and follows the conventional secretory pathway to be released into the extracellular compartment without the involvement of proteolysis. This particular secretion mechanism yields a soluble TM536, which is poorly modified by chondroitin sulfate glycosaminoglycan compared with conventionally secreted soluble forms of TM, and therefore has a suboptimal capacity to mediate thrombin-dependent activation of protein C (PC). We also showed that TM536 cellular trafficking was altered, with retention in the early secretory pathway and increased sensitivity to ER-associated degradation. As expected, activation of ER-associated degradation increased TM536 degradation and reduced its release. The expression of TM536 at the cell surface was low, and its distribution in lipid raft-like membrane microdomains was altered, resulting in low thrombin-dependent PC activation on the cell surface.
© 2024 by The American Society of Hematology. Licensed under Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International (CC BY-NC-ND 4.0), permitting only noncommercial, nonderivative use with attribution. All other rights reserved.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures





References
-
- Loghmani H, Conway EM. Exploring traditional and nontraditional roles for thrombomodulin. Blood. 2018;132(2):148–158. - PubMed
-
- Mulder AB, Smit JW, Bom VJ, Blom NR, Halie MR, van der Meer J. Association of endothelial tissue factor and thrombomodulin with caveolae. Blood. 1996;88(9):3667–3670. - PubMed
-
- Chen P-K, Chang B-I, Kuo C-H, et al. Thrombomodulin functions as a plasminogen receptor to modulate angiogenesis. FASEB J. 2013;27(11):4520–4531. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
