

Adipocyte deletion of the oxygen-sensor PHD2 sustains elevated energy expenditure at thermoneutrality
- PMID: 39209825
- PMCID: PMC11362468
- DOI: 10.1038/s41467-024-51718-7
Adipocyte deletion of the oxygen-sensor PHD2 sustains elevated energy expenditure at thermoneutrality
Abstract
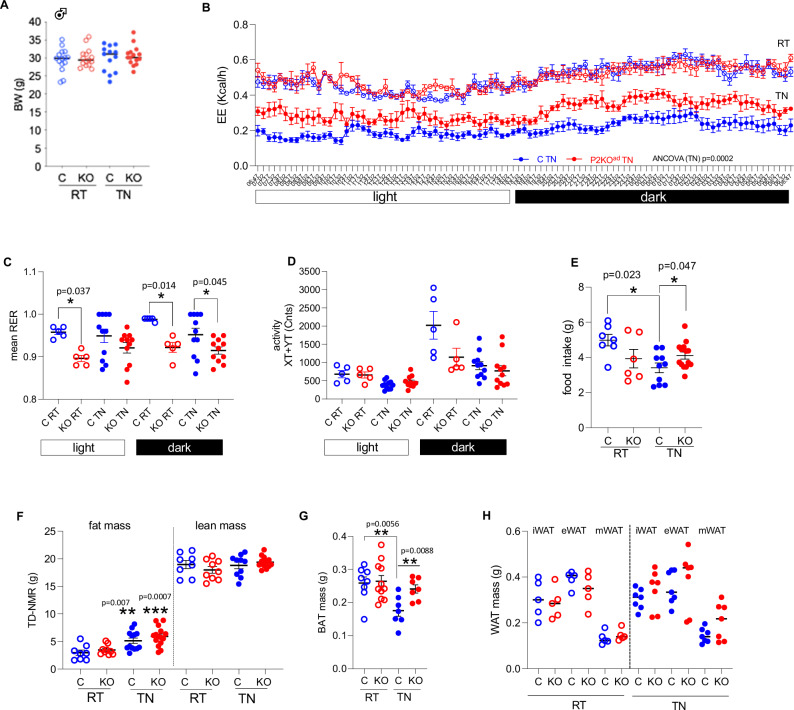
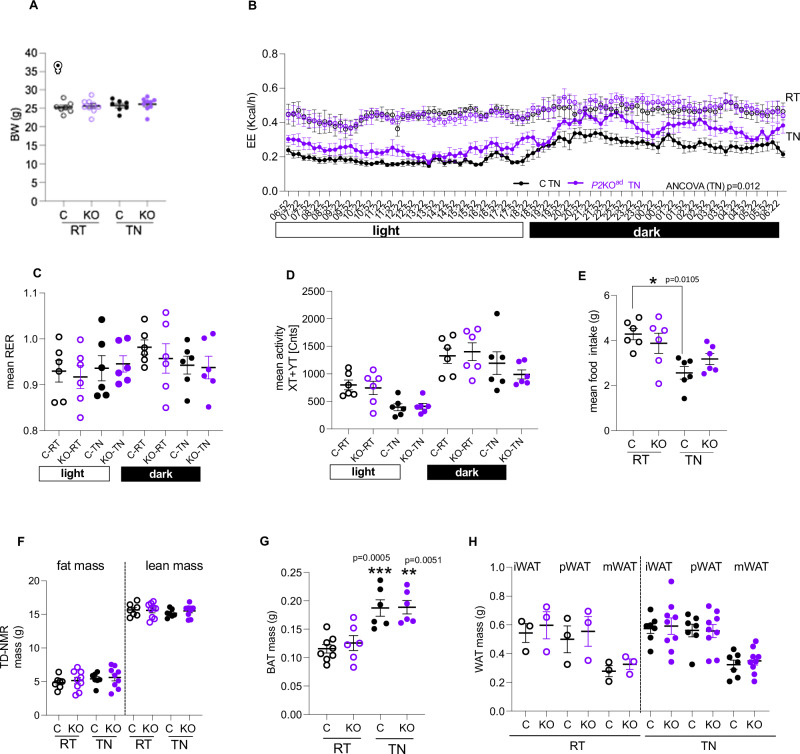
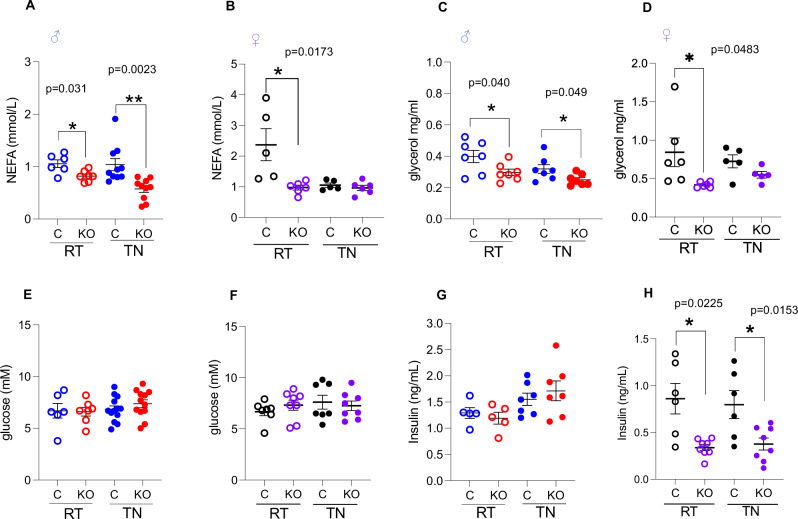
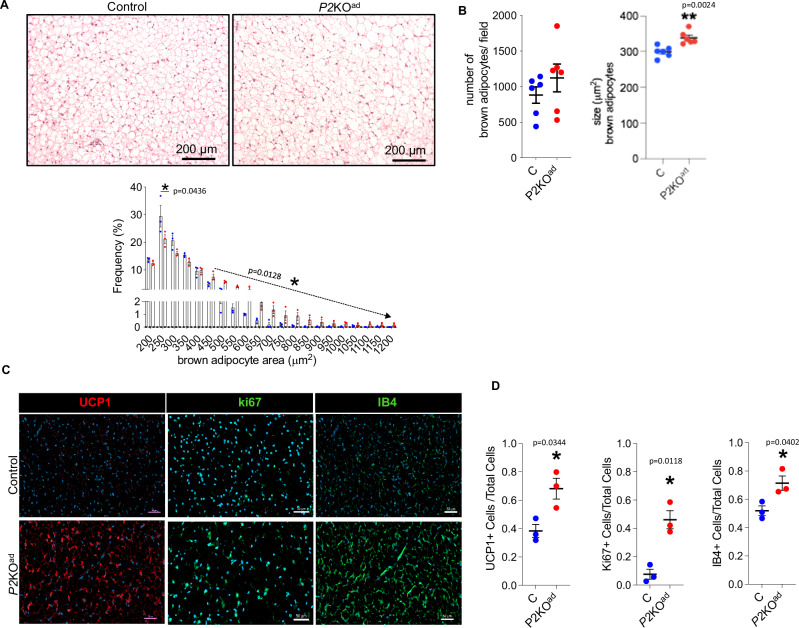
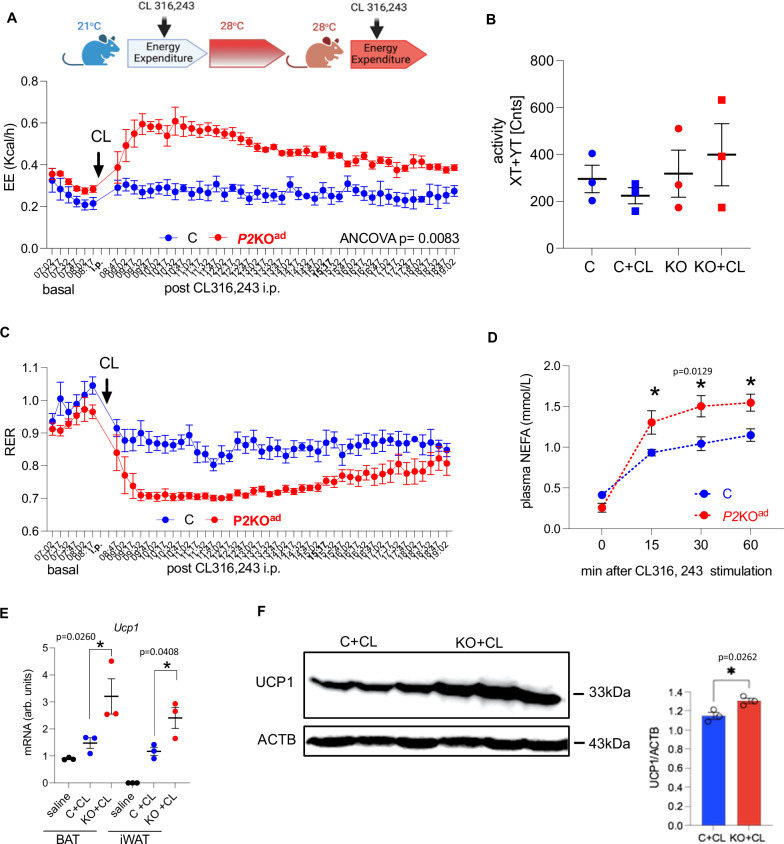
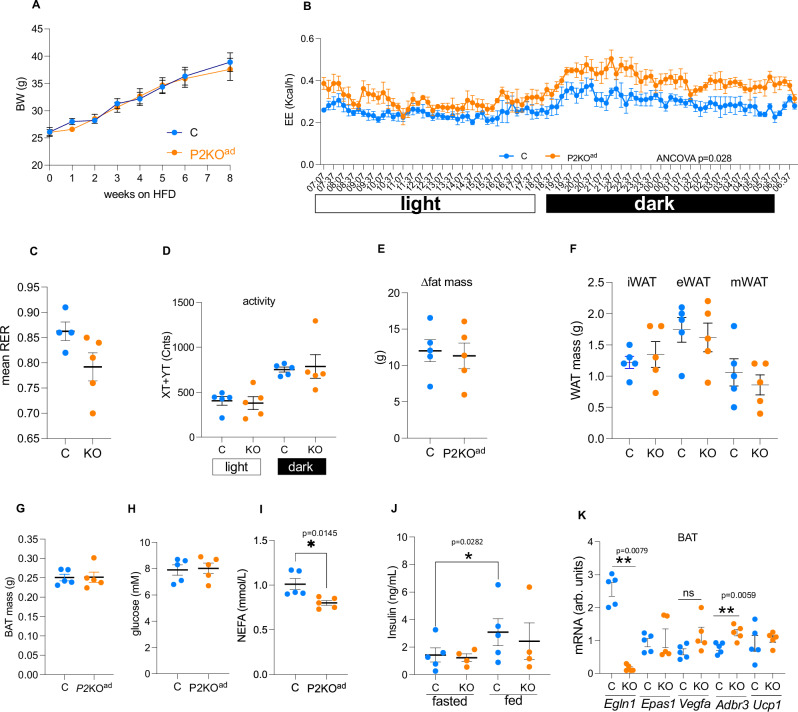
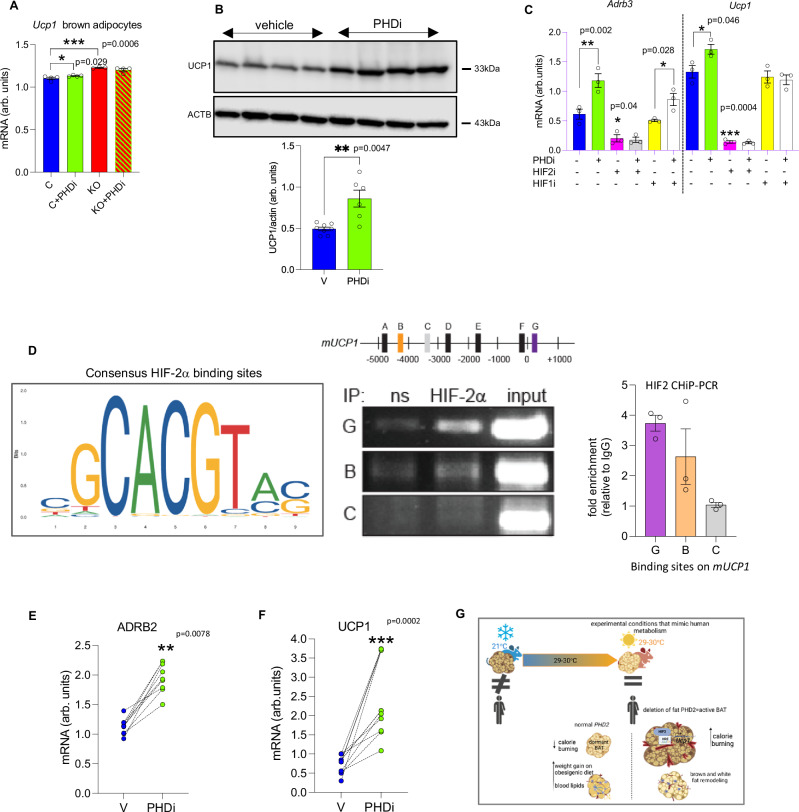
Enhancing thermogenic brown adipose tissue (BAT) function is a promising therapeutic strategy for metabolic disease. However, predominantly thermoneutral modern human living conditions deactivate BAT. We demonstrate that selective adipocyte deficiency of the oxygen-sensor HIF-prolyl hydroxylase (PHD2) gene overcomes BAT dormancy at thermoneutrality. Adipocyte-PHD2-deficient mice maintain higher energy expenditure having greater BAT thermogenic capacity. In human and murine adipocytes, a PHD inhibitor increases Ucp1 levels. In murine brown adipocytes, antagonising the major PHD2 target, hypoxia-inducible factor-(HIF)-2a abolishes Ucp1 that cannot be rescued by PHD inhibition. Mechanistically, PHD2 deficiency leads to HIF2 stabilisation and binding of HIF2 to the Ucp1 promoter, thus enhancing its expression in brown adipocytes. Serum proteomics analysis of 5457 participants in the deeply phenotyped Age, Gene and Environment Study reveal that serum PHD2 associates with increased risk of metabolic disease. Here we show that adipose-PHD2-inhibition is a therapeutic strategy for metabolic disease and identify serum PHD2 as a disease biomarker.
© 2024. The Author(s).
Conflict of interest statement
L.L.J. is an employee and stockholder of Novartis. V.E. V.G. C.J.S., N.M.M, R.H.S, M.G.S, T.C., R.G.M., I.P.C, K.F., R.W., C.B., M.C., A.C., T.J.S, S.G., M.M.P., I.K., S.G., T.J.S., and Z.M. declare no competing interests.
Figures
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
