

YY1-controlled regulatory connectivity and transcription are influenced by the cell cycle
- PMID: 39210046
- PMCID: PMC11687402
- DOI: 10.1038/s41588-024-01871-y
YY1-controlled regulatory connectivity and transcription are influenced by the cell cycle
Abstract
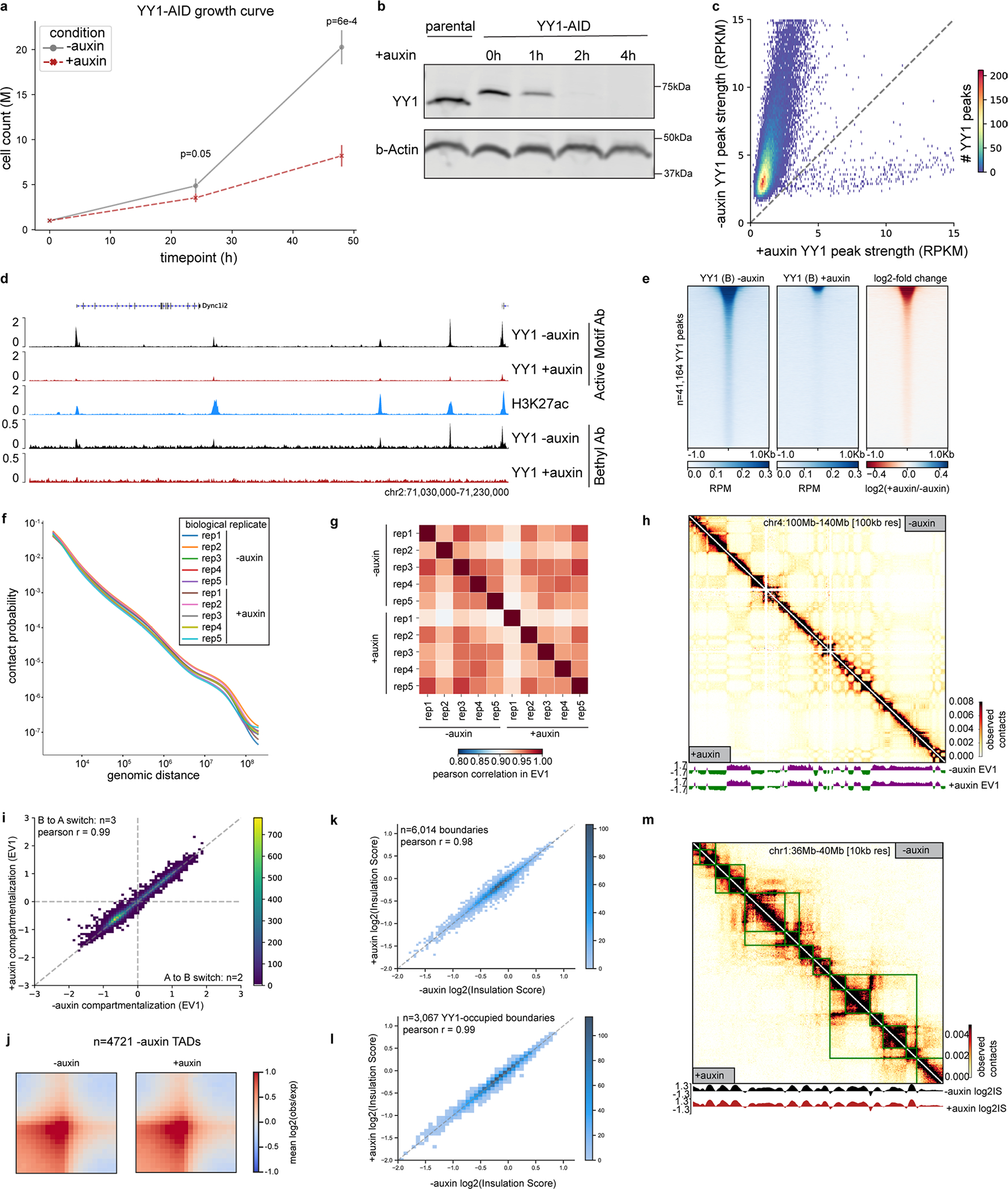
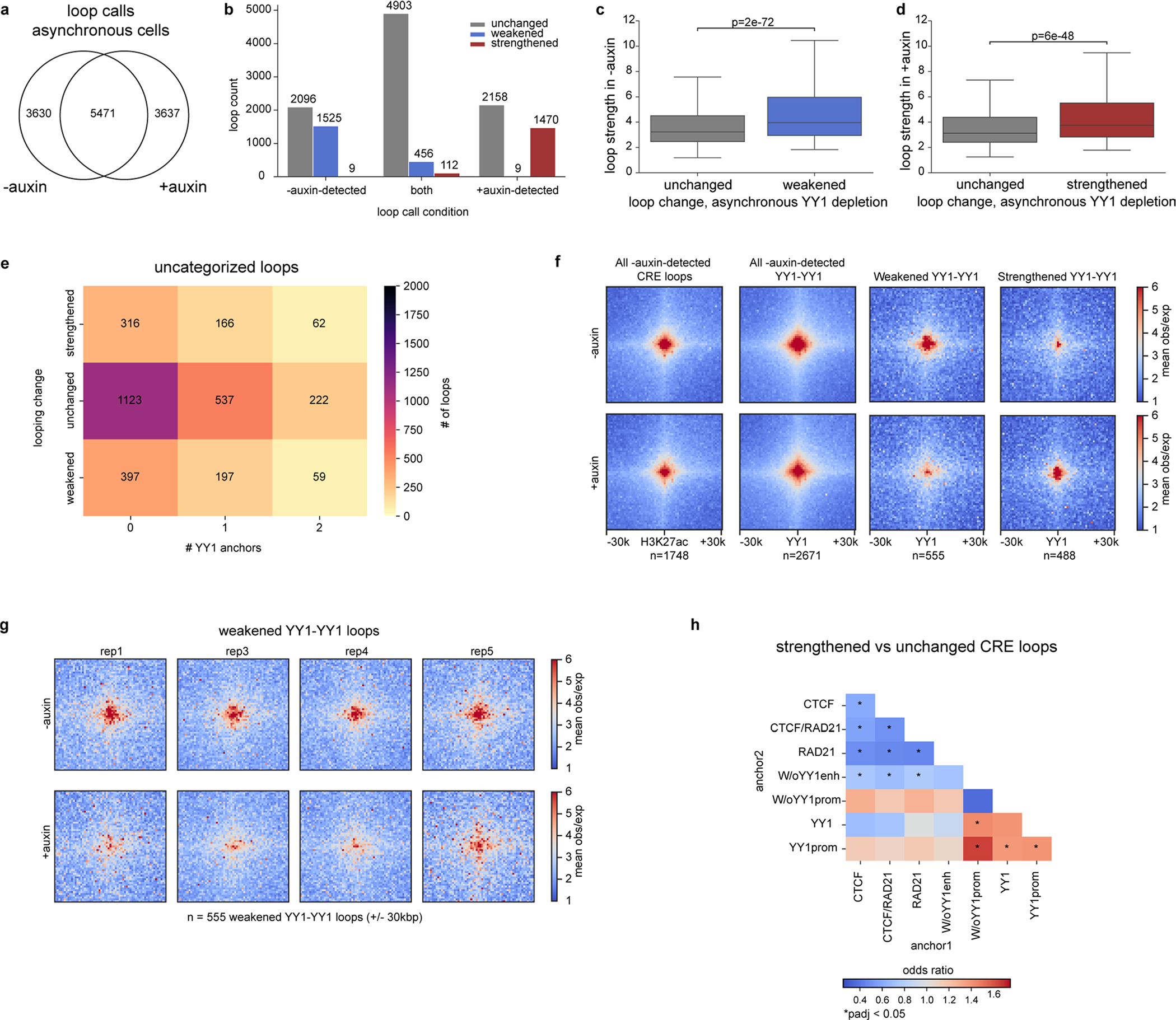
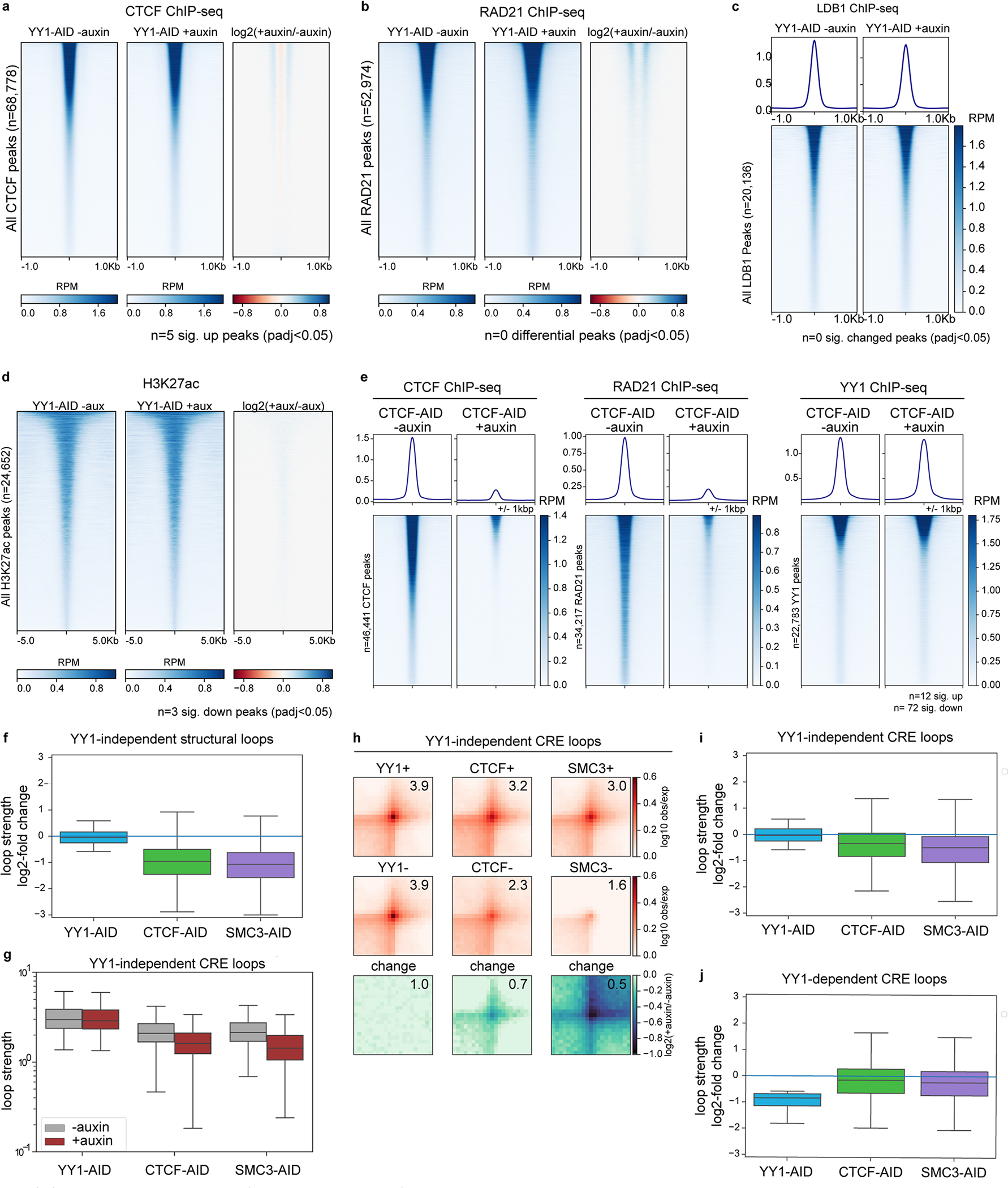
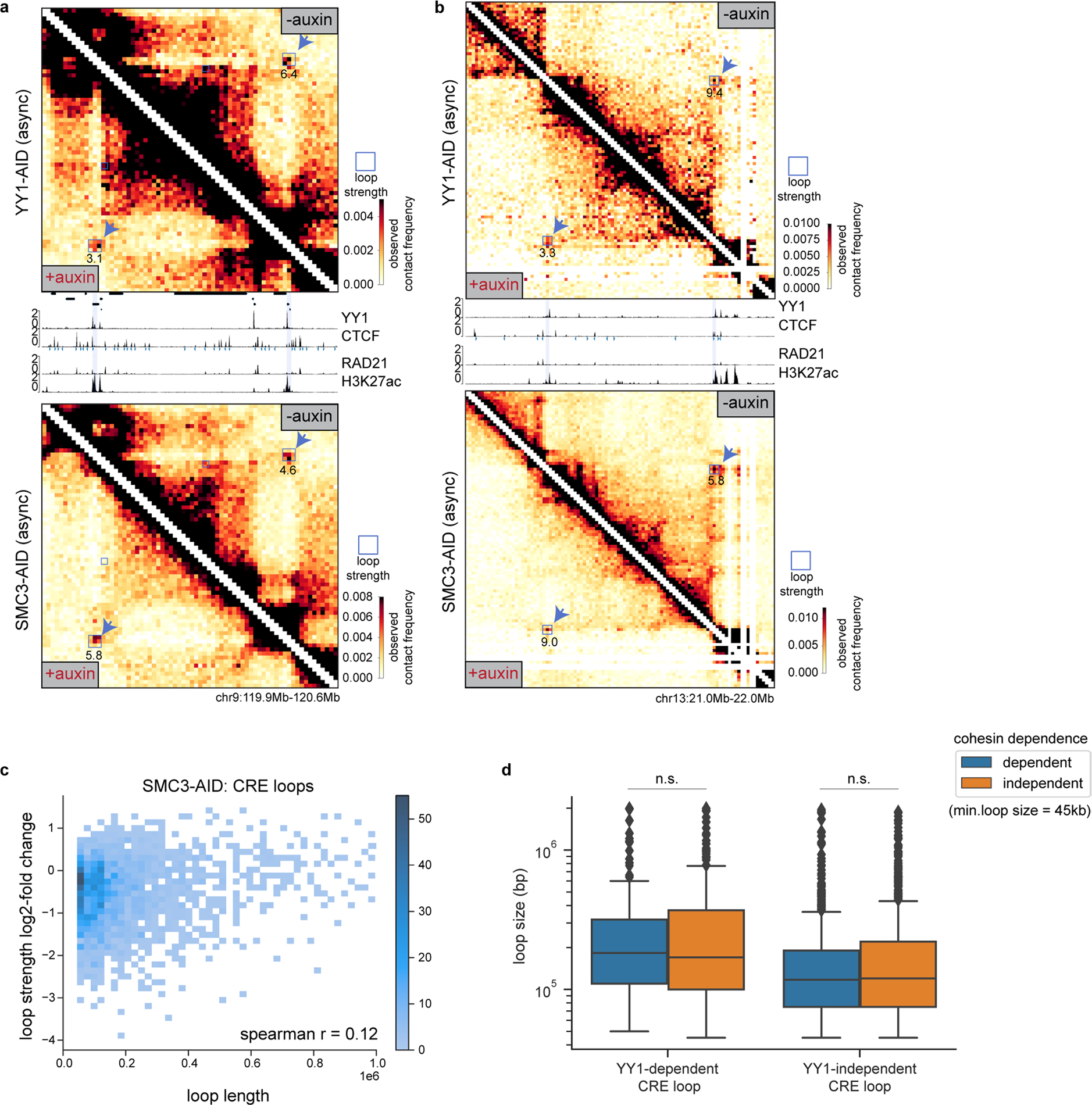
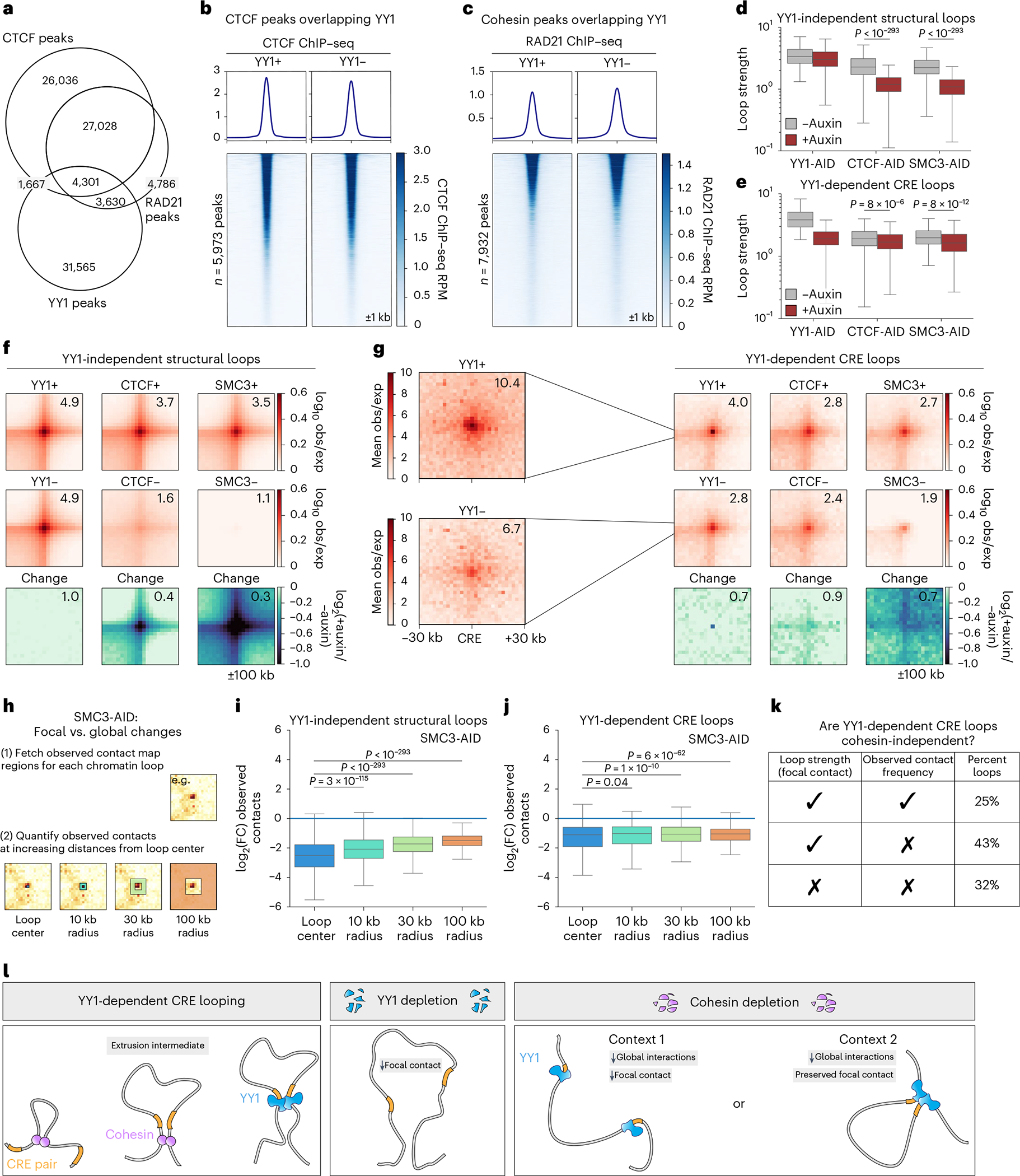
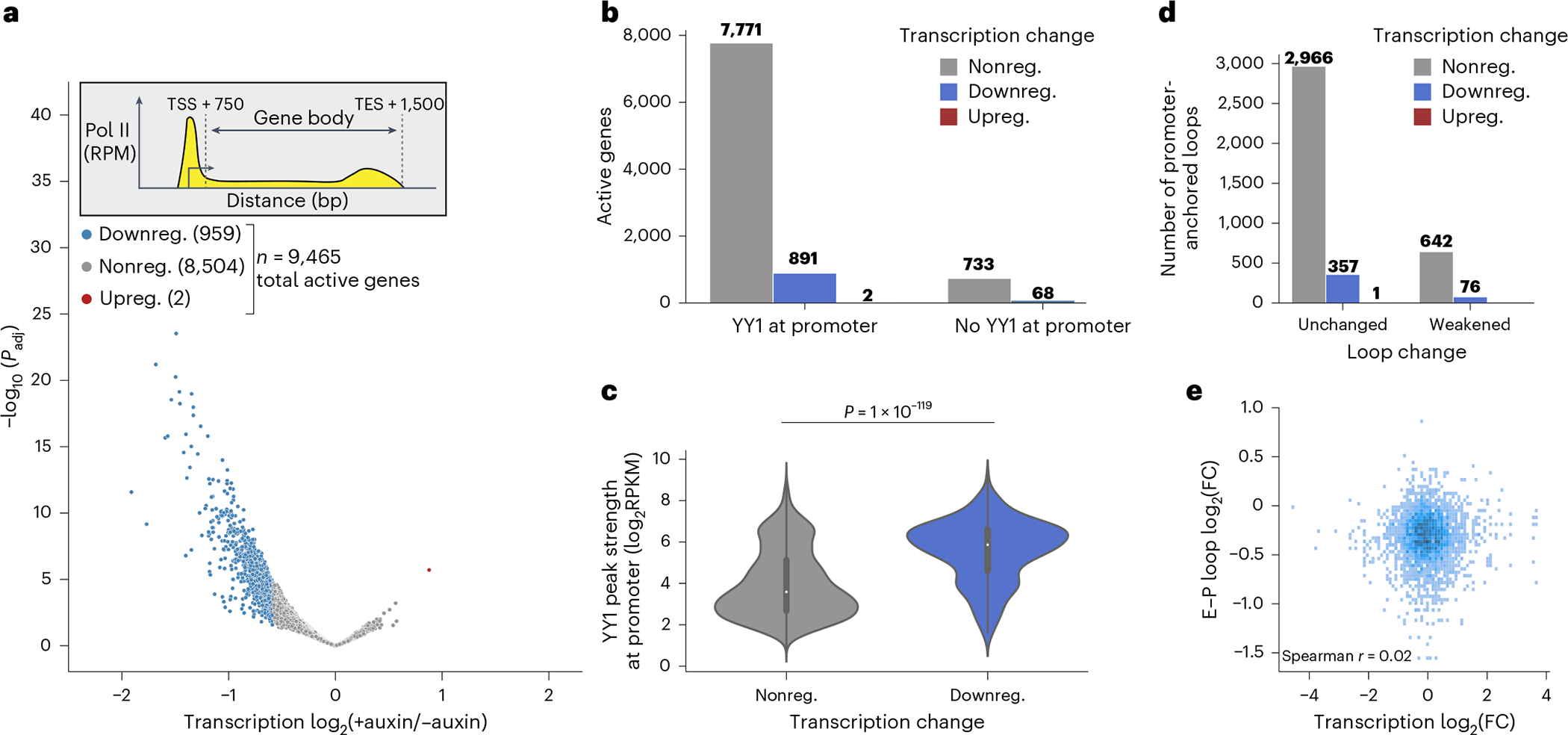
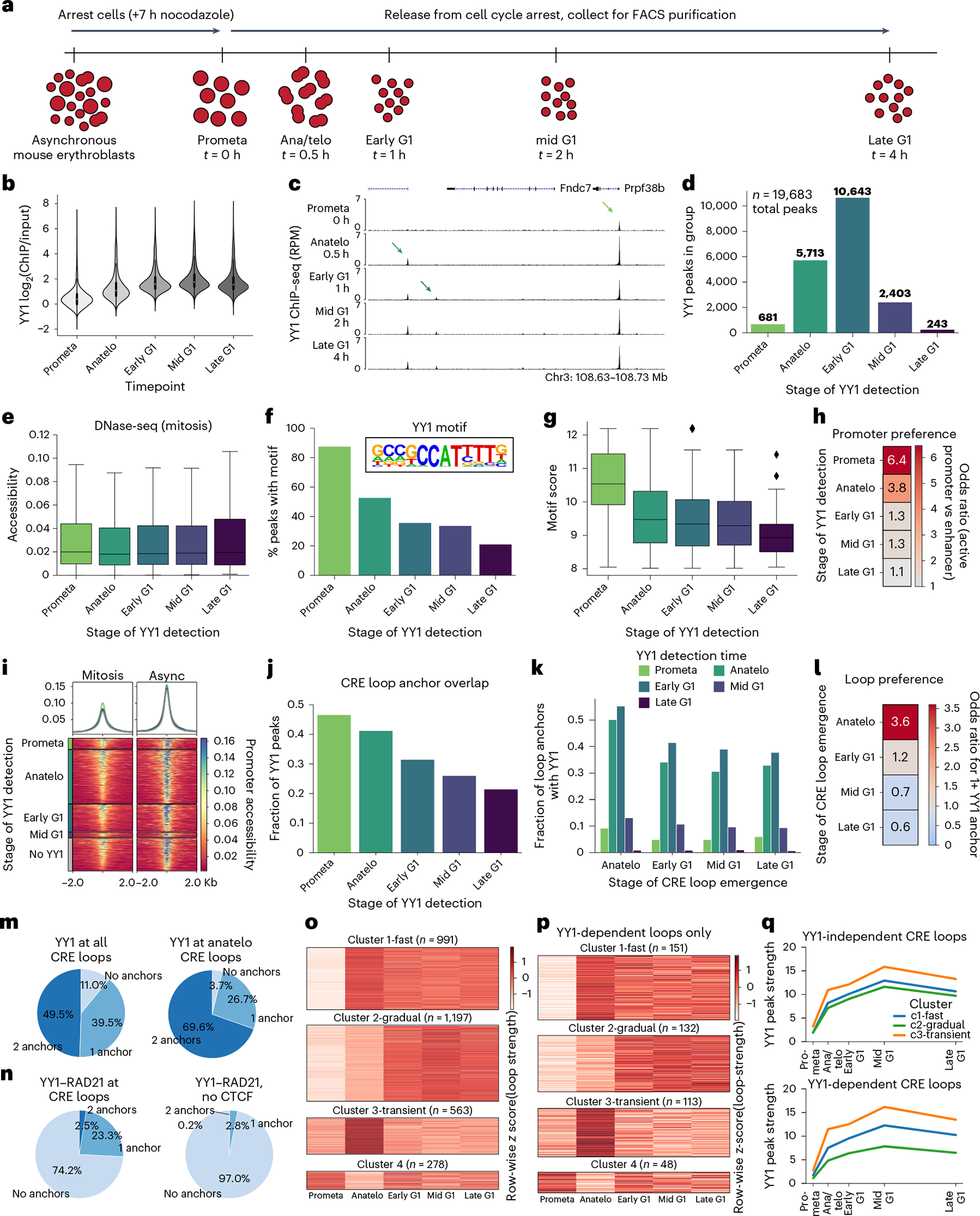
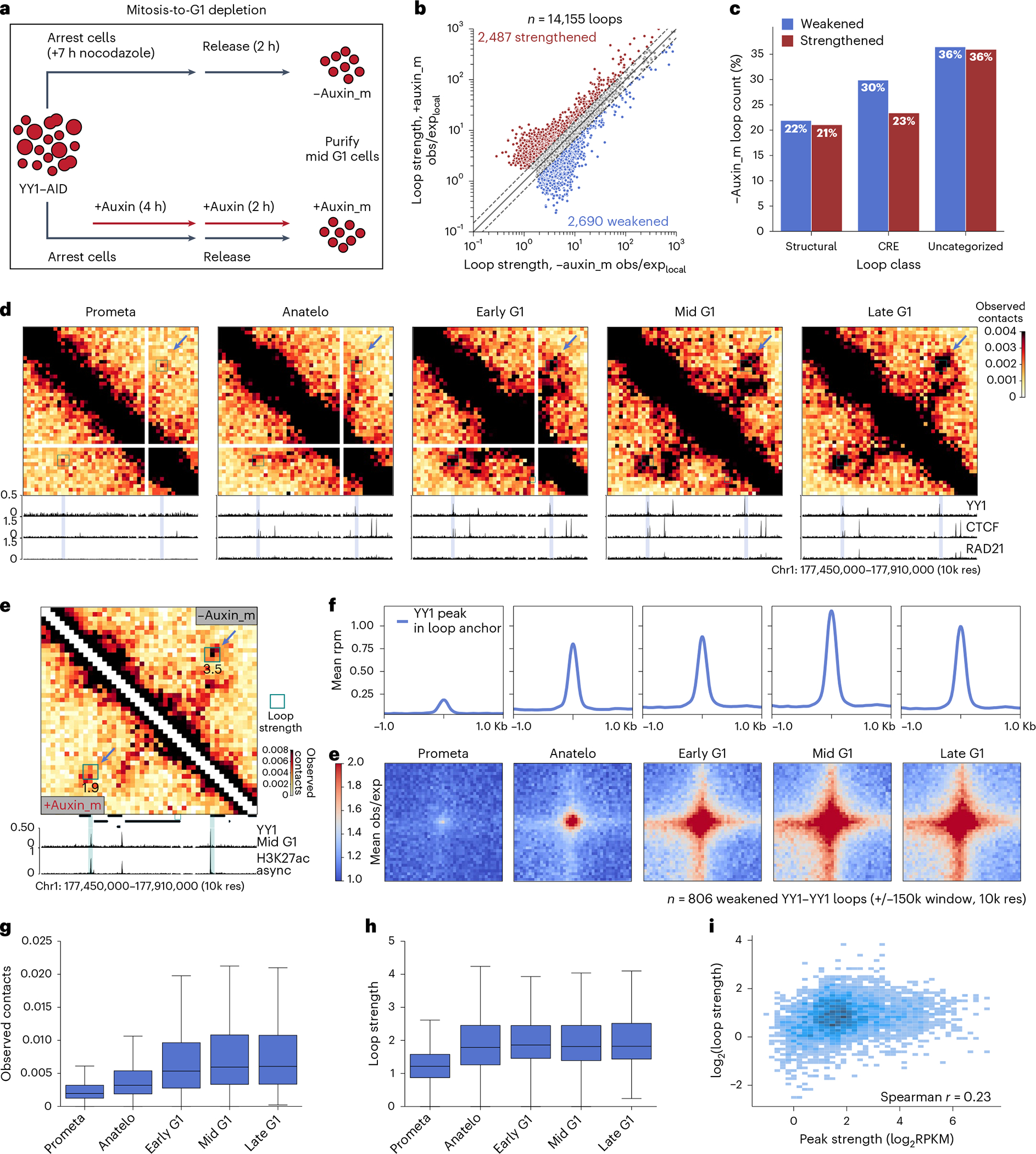
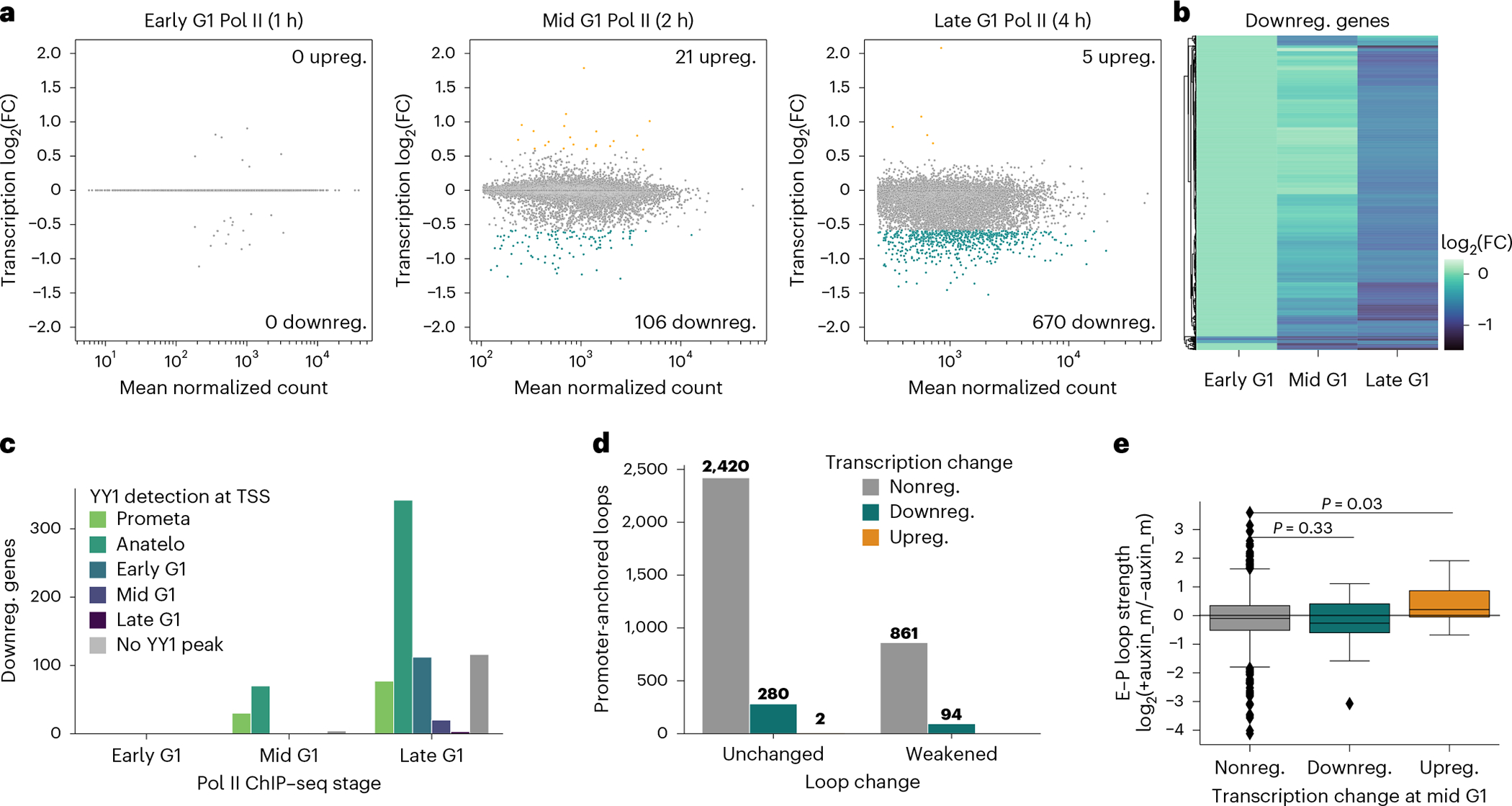
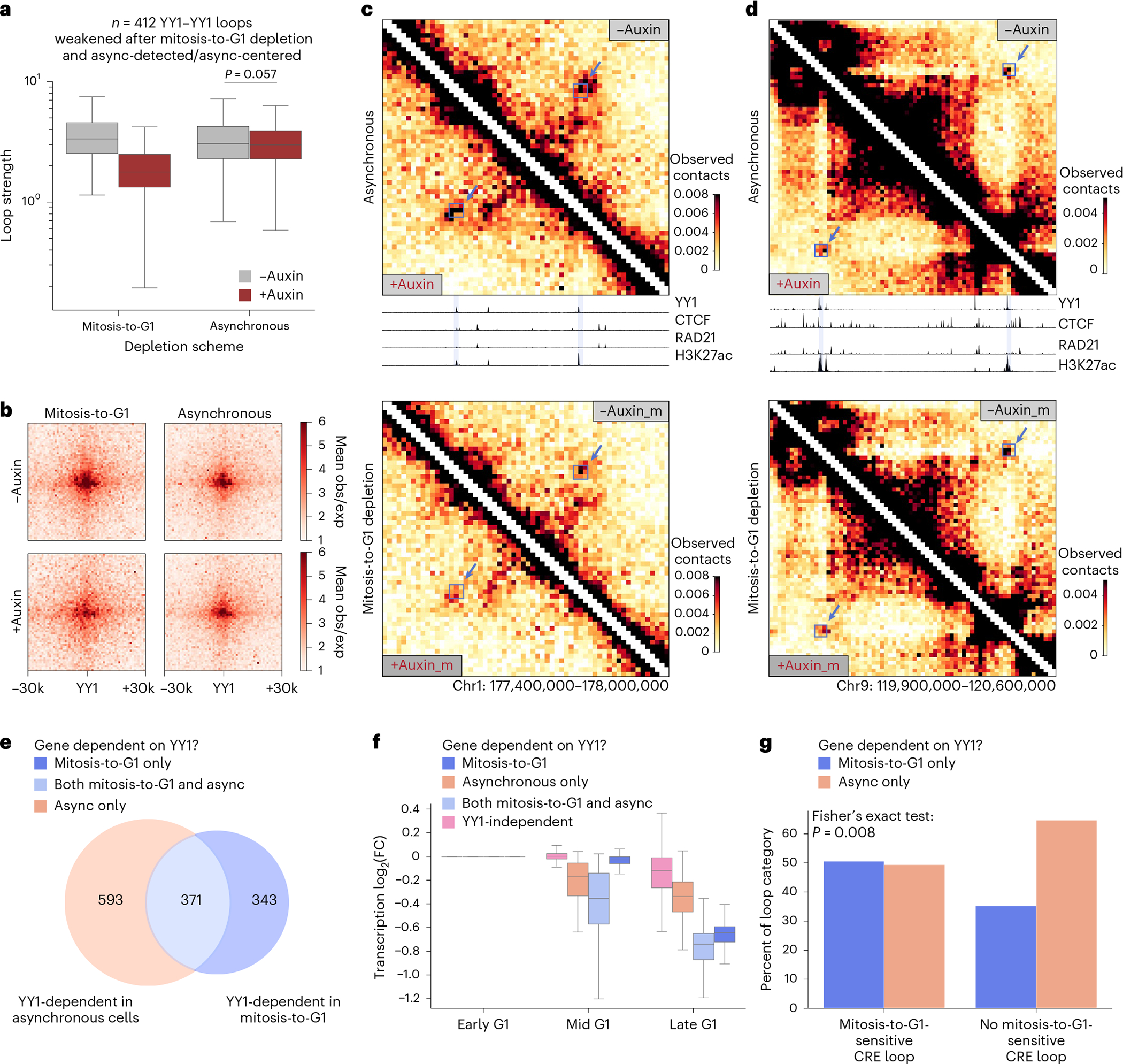
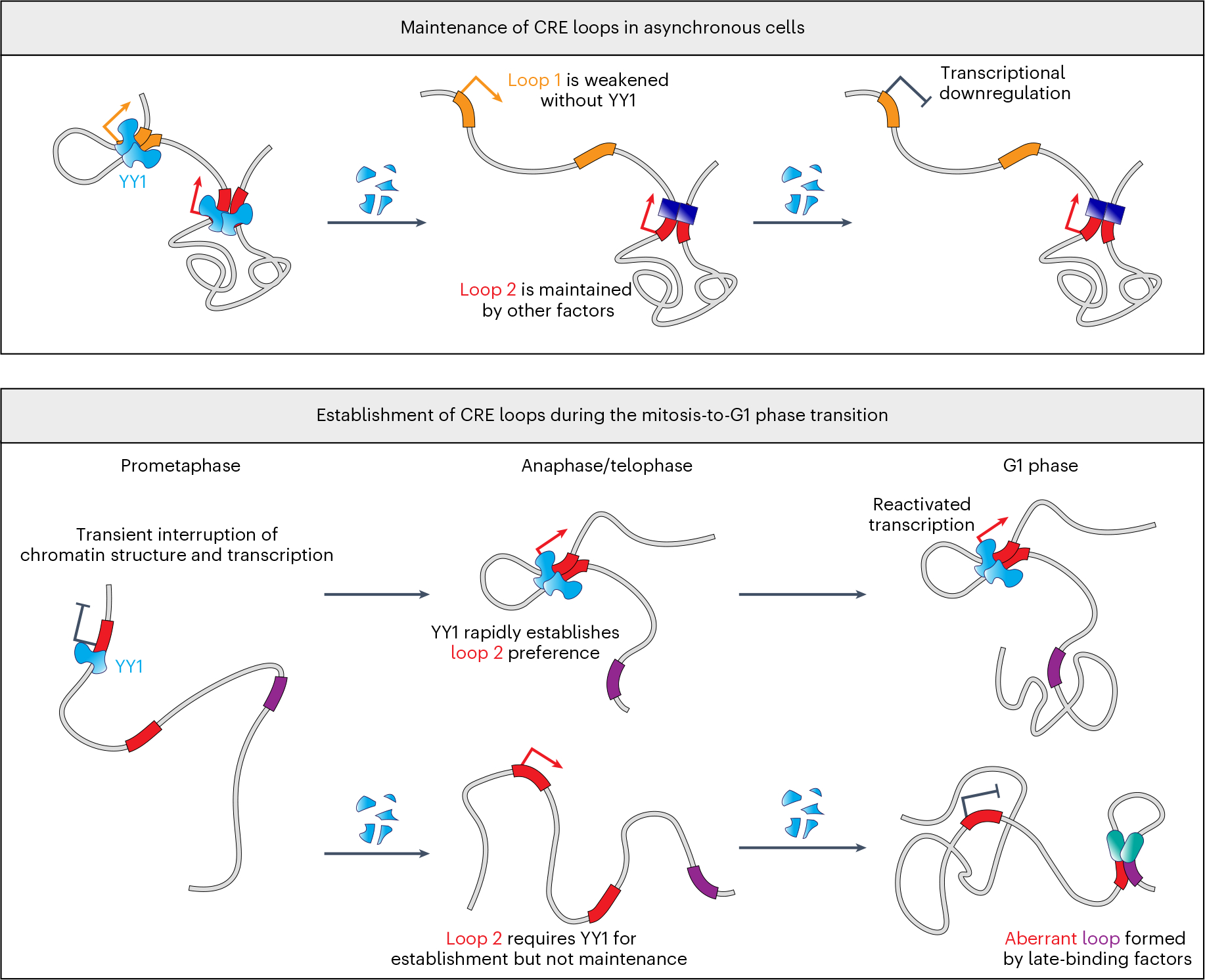
Few transcription factors have been examined for their direct roles in physically connecting enhancers and promoters. Here acute degradation of Yin Yang 1 (YY1) in erythroid cells revealed its requirement for the maintenance of numerous enhancer-promoter loops, but not compartments or domains. Despite its reported ability to interact with cohesin, the formation of YY1-dependent enhancer-promoter loops does not involve stalling of cohesin-mediated loop extrusion. Integrating mitosis-to-G1-phase dynamics, we observed partial retention of YY1 on mitotic chromatin, predominantly at gene promoters, followed by rapid rebinding during mitotic exit, coinciding with enhancer-promoter loop establishment. YY1 degradation during the mitosis-to-G1-phase interval revealed a set of enhancer-promoter loops that require YY1 for establishment during G1-phase entry but not for maintenance in interphase, suggesting that cell cycle stage influences YY1's architectural function. Thus, as revealed here for YY1, chromatin architectural functions of transcription factors can vary in their interplay with CTCF and cohesin as well as by cell cycle stage.
© 2024. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing interests
The authors declare no competing interests.
Figures
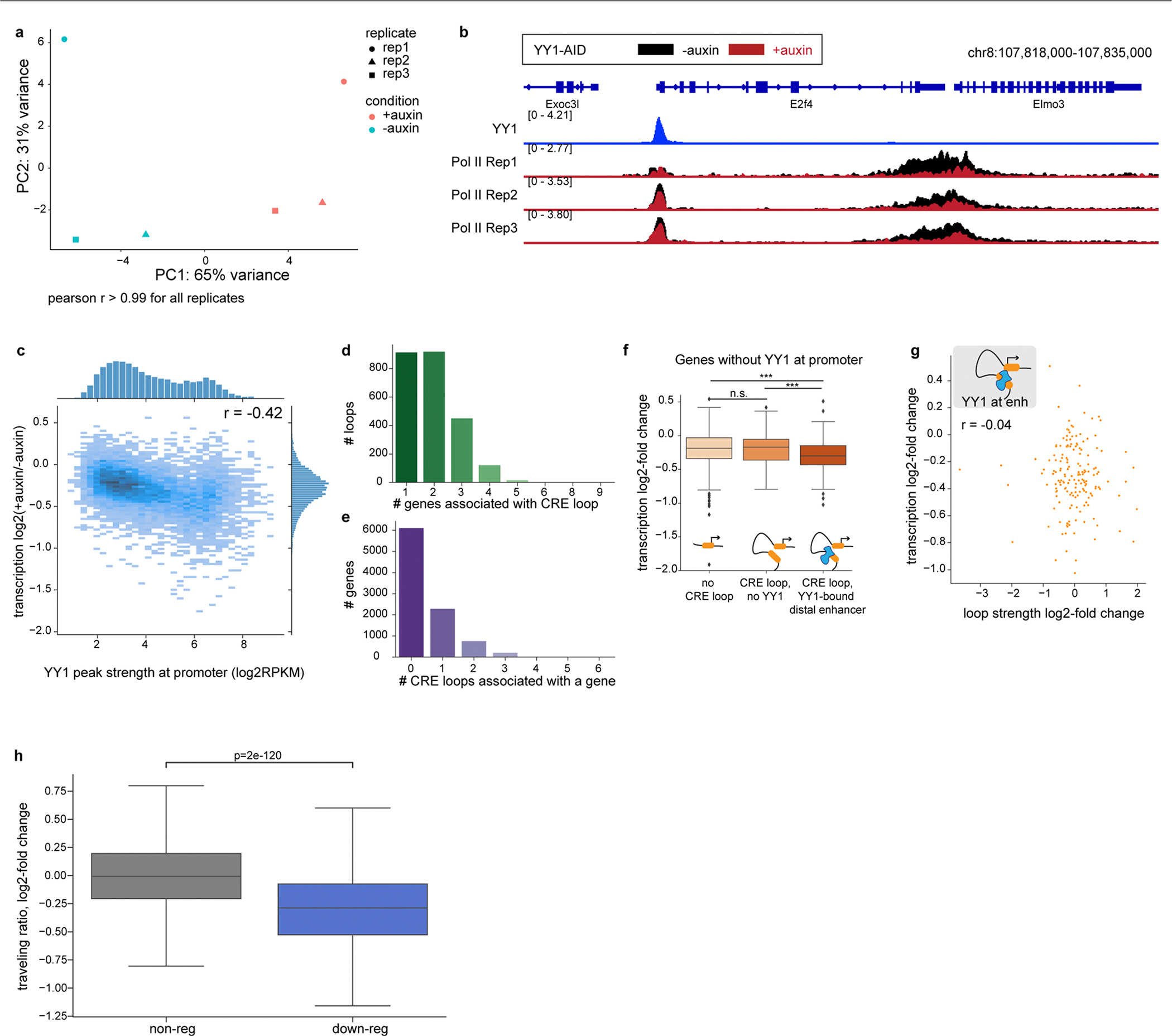
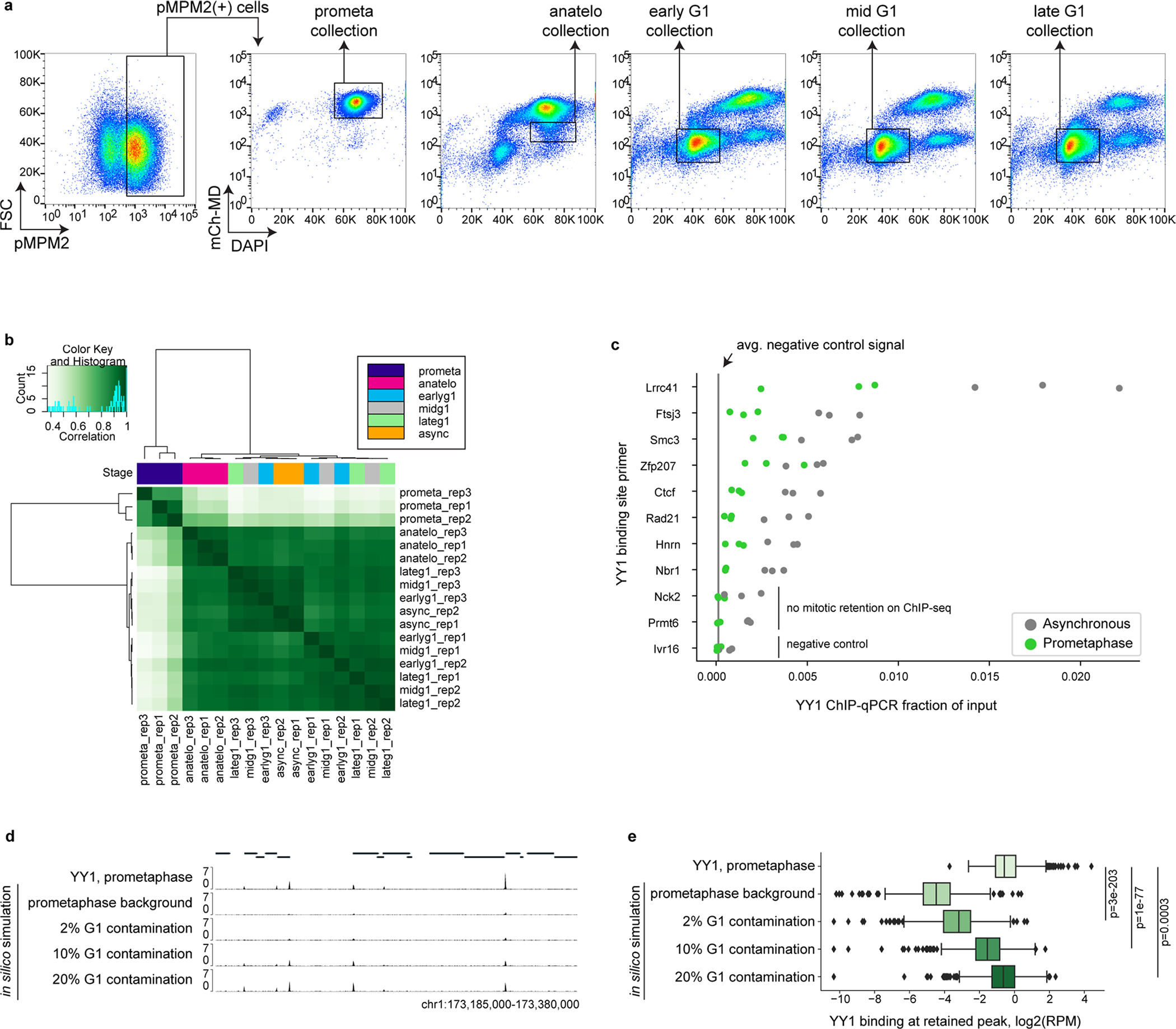
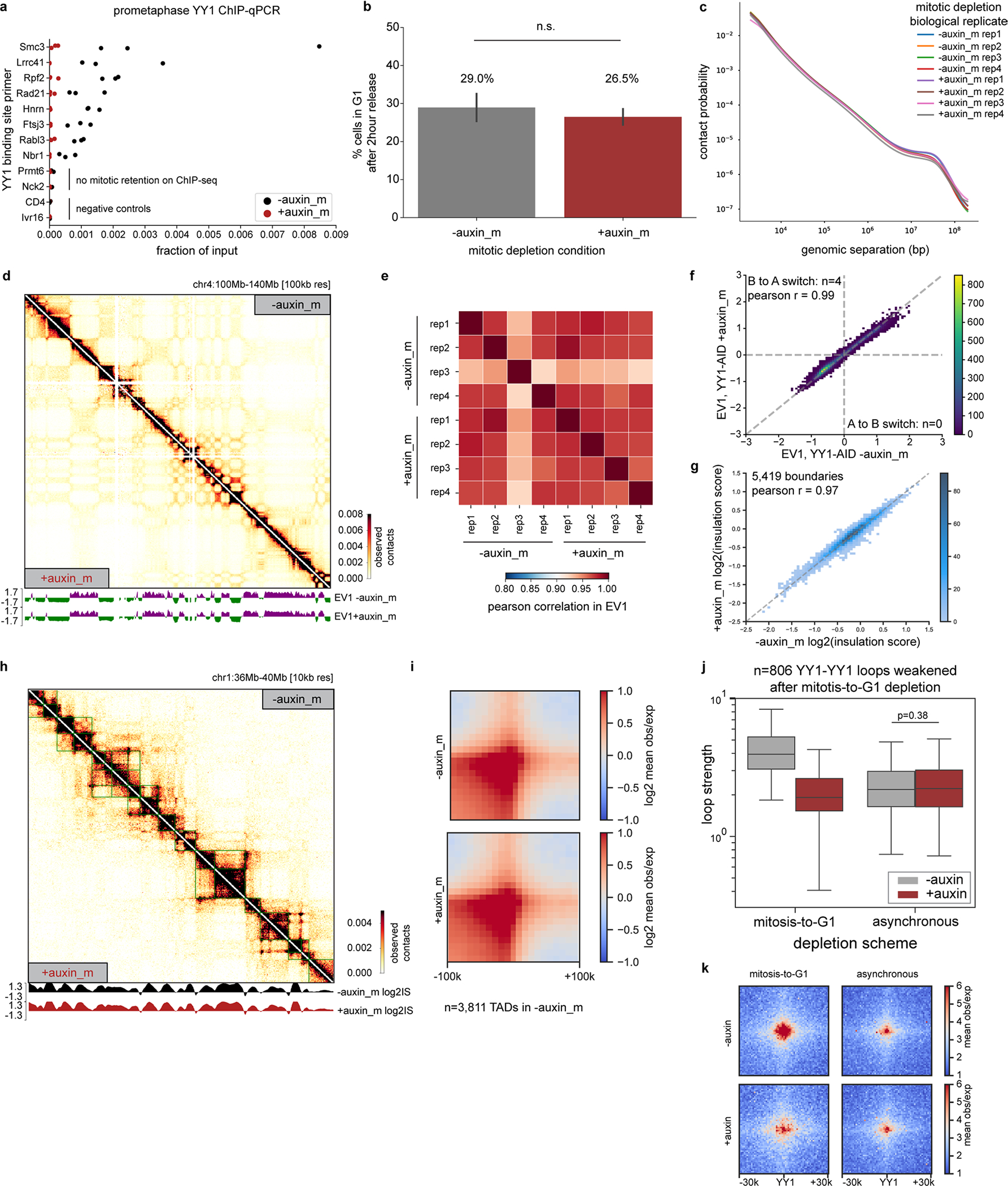
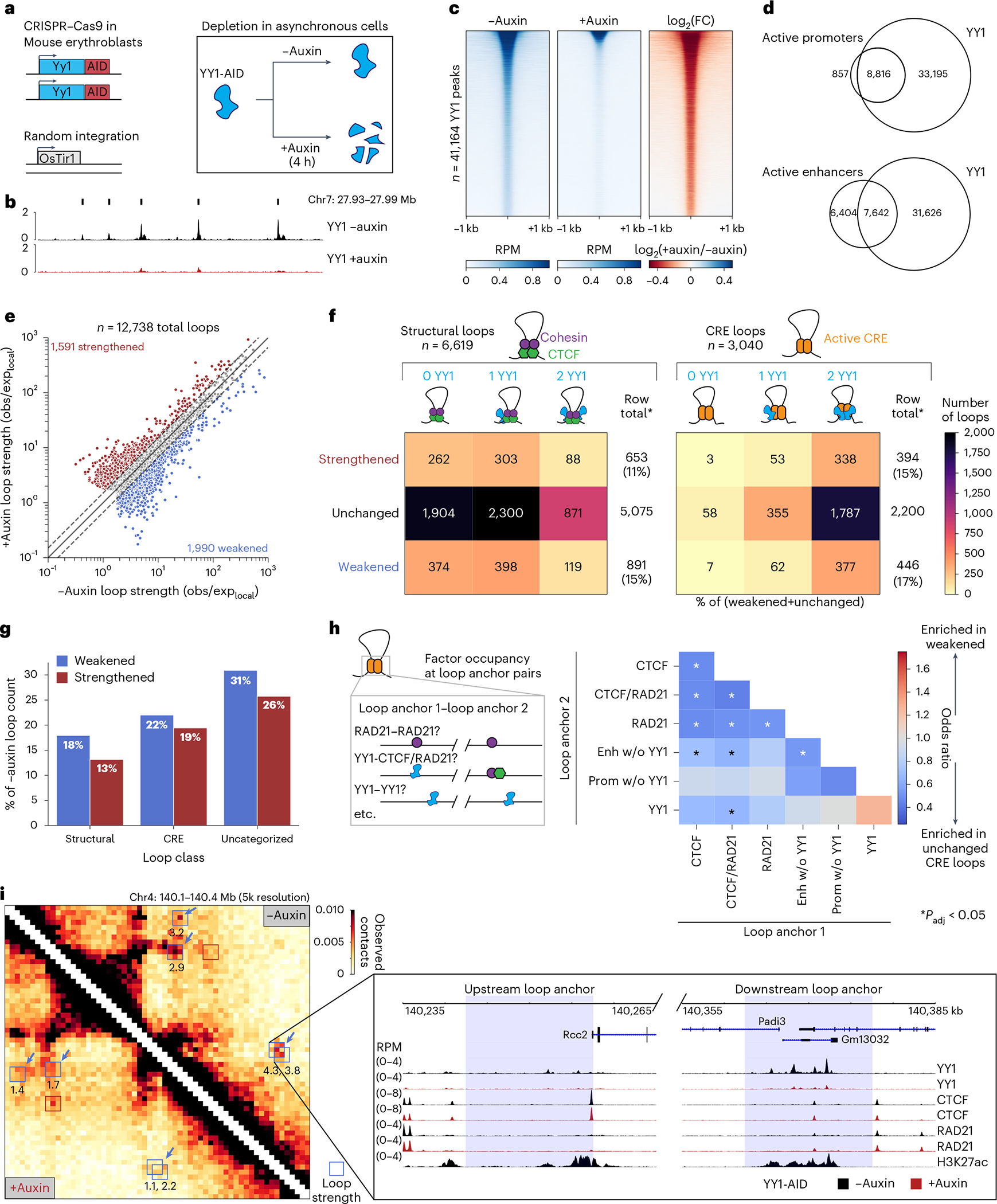
Similar articles
-
LDB1 establishes multi-enhancer networks to regulate gene expression.Mol Cell. 2025 Jan 16;85(2):376-393.e9. doi: 10.1016/j.molcel.2024.11.037. Epub 2024 Dec 24. Mol Cell. 2025. PMID: 39721581
-
Prediction of YY1 loop anchor based on multi-omics features.Methods. 2024 Dec;232:96-106. doi: 10.1016/j.ymeth.2024.11.004. Epub 2024 Nov 7. Methods. 2024. PMID: 39521361
-
Prediction of cell-type-specific cohesin-mediated chromatin loops based on chromatin state.Methods. 2024 Jun;226:151-160. doi: 10.1016/j.ymeth.2024.04.014. Epub 2024 Apr 24. Methods. 2024. PMID: 38670416
-
Beyond genomic weaving: molecular roles for CTCF outside cohesin loop extrusion.Curr Opin Genet Dev. 2025 Feb;90:102298. doi: 10.1016/j.gde.2024.102298. Epub 2024 Dec 21. Curr Opin Genet Dev. 2025. PMID: 39709822 Free PMC article. Review.
-
Chromatin insulator mechanisms ensure accurate gene expression by controlling overall 3D genome organization.Curr Opin Genet Dev. 2024 Aug;87:102208. doi: 10.1016/j.gde.2024.102208. Epub 2024 May 28. Curr Opin Genet Dev. 2024. PMID: 38810546 Free PMC article. Review.
Cited by
-
Putative looping factor ZNF143/ZFP143 is an essential transcriptional regulator with no looping function.Mol Cell. 2025 Jan 2;85(1):9-23.e9. doi: 10.1016/j.molcel.2024.11.032. Epub 2024 Dec 20. Mol Cell. 2025. PMID: 39708803 Free PMC article.
-
Molecular mechanisms altering cell identity in cancer.Oncogene. 2025 Jul;44(26):2117-2126. doi: 10.1038/s41388-025-03314-2. Epub 2025 Feb 26. Oncogene. 2025. PMID: 40011573 Review.
-
Klf5-adjacent super-enhancer functions as a 3D genome structure-dependent transcriptional driver to safeguard ESC identity.Nat Commun. 2025 Jul 1;16(1):5540. doi: 10.1038/s41467-025-60389-x. Nat Commun. 2025. PMID: 40592854 Free PMC article.
-
Dynamics of microcompartment formation at the mitosis-to-G1 transition.bioRxiv [Preprint]. 2024 Sep 16:2024.09.16.611917. doi: 10.1101/2024.09.16.611917. bioRxiv. 2024. PMID: 39345388 Free PMC article. Preprint.
-
LDB1 establishes multi-enhancer networks to regulate gene expression.Mol Cell. 2025 Jan 16;85(2):376-393.e9. doi: 10.1016/j.molcel.2024.11.037. Epub 2024 Dec 24. Mol Cell. 2025. PMID: 39721581
References
MeSH terms
Substances
Grants and funding
- R24DK106766/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- T32HG000046/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- S06 GM008216/GM/NIGMS NIH HHS/United States
- T32GM008216/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R01 DK058044/DK/NIDDK NIH HHS/United States
- R01DK054937/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- T32 GM007170/GM/NIGMS NIH HHS/United States
- R01 DK054937/DK/NIDDK NIH HHS/United States
- U01 DK127405/DK/NIDDK NIH HHS/United States
- T32 HG000046/HG/NHGRI NIH HHS/United States
- 32100422/National Natural Science Foundation of China (National Science Foundation of China)
- F30 DK132824/DK/NIDDK NIH HHS/United States
- T32 GM008216/GM/NIGMS NIH HHS/United States
- U01DK127405/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- T32GM007170/U.S. Department of Health & Human Services | NIH | National Institute of General Medical Sciences (NIGMS)
- R24 DK106766/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
