

Recycling machinery of integrin coupled with focal adhesion turnover via RAB11-UNC13D-FAK axis for migration of pancreatic cancer cells
- PMID: 39210440
- PMCID: PMC11360766
- DOI: 10.1186/s12967-024-05630-9
Recycling machinery of integrin coupled with focal adhesion turnover via RAB11-UNC13D-FAK axis for migration of pancreatic cancer cells
Abstract
Background: Recycling of integrin via endosomal vesicles is critical for the migration of cancer cells, which leads to the metastasis of pancreatic cancer and devastating cancer-related death. So, new diagnostic and therapeutic molecules which target the recycling of endosomal vesicles need to be developed.
Methods: Public databases including TCGA, ICGC, GSE21501, GSE28735, and GENT are analyzed to derive diagnostic and therapeutic targets. To reveal biological roles and underlying mechanisms of molecular targets, various molecular biological experiments were conducted.
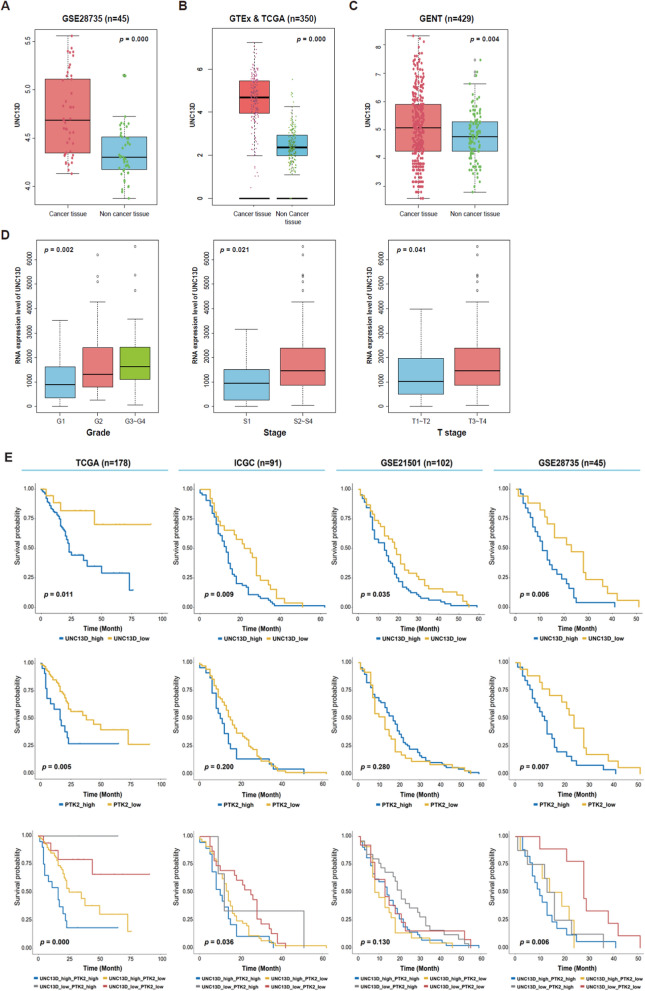
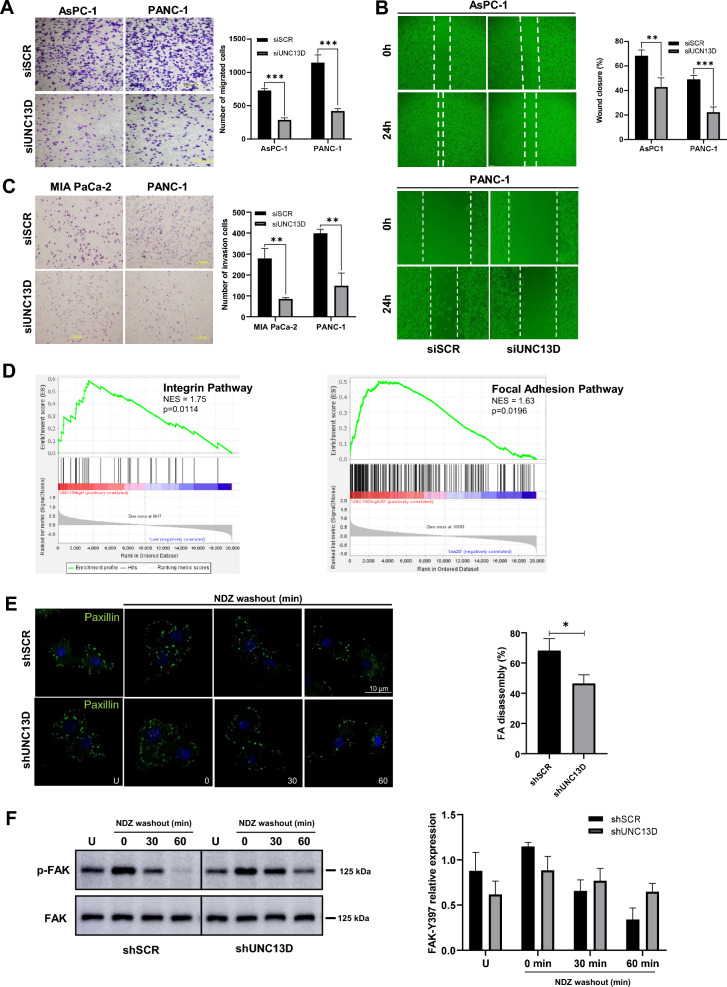
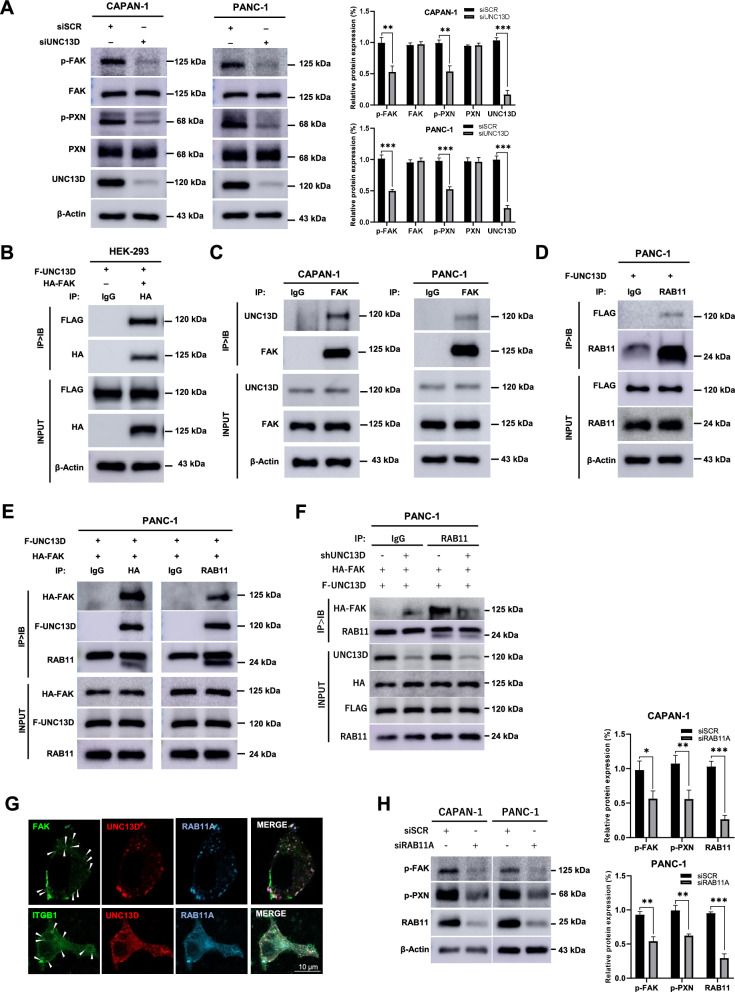
Results: First, we identified UNC13D's overexpression in patients with pancreatic cancer (n = 824) and its prognostic significance and high hazard ratio (HR) in four independent pancreatic cancer cohorts (TCGA, n = 178, p = 0.014, HR = 3.629; ICGC, n = 91, p = 0.000, HR = 4.362; GSE21501, n = 102, p = 0.002, HR = 2.339; GSE28735, n = 45, p = 0.022, HR = 2.681). Additionally, its expression is associated with the clinicopathological progression of pancreatic cancer. Further biological studies have shown that UNC13D regulates the migration of pancreatic cancer cells by coupling the exocytosis of recycling endosomes with focal adhesion turnover via the regulation of FAK phosphorylation. Immunoprecipitation and immunocytochemistry showed the formation of the RAB11-UNC13D-FAK axis in endosomes during integrin recycling. We observed that UNC13D directly interacted with the FERM domain of FAK and regulated FAK phosphorylation in a calcium-dependent manner. Finally, we found co-expression of UNC13D and FAK showed the poorest survival (TCGA, p = 0.000; ICGC, p = 0.036; GSE28735, p = 0.006).
Conclusions: We highlight that UNC13D, a novel prognostic factor, promotes pancreatic cancer progression by coupling integrin recycling with focal adhesion turnover via the RAB11-UNC13D-FAK axis for the migration of pancreatic cancer cells.
Keywords: FAK; Focal adhesion; Integrin; Migration; Pancreatic cancer; Prognosis; RAB11; UNC13D.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no potential conflicts of interest.
Figures


References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
