

Metabolic immaturity and breastmilk bile acid metabolites are central determinants of heightened newborn vulnerability to norovirus diarrhea
- PMID: 39214086
- PMCID: PMC11392616
- DOI: 10.1016/j.chom.2024.08.003
Metabolic immaturity and breastmilk bile acid metabolites are central determinants of heightened newborn vulnerability to norovirus diarrhea
Abstract
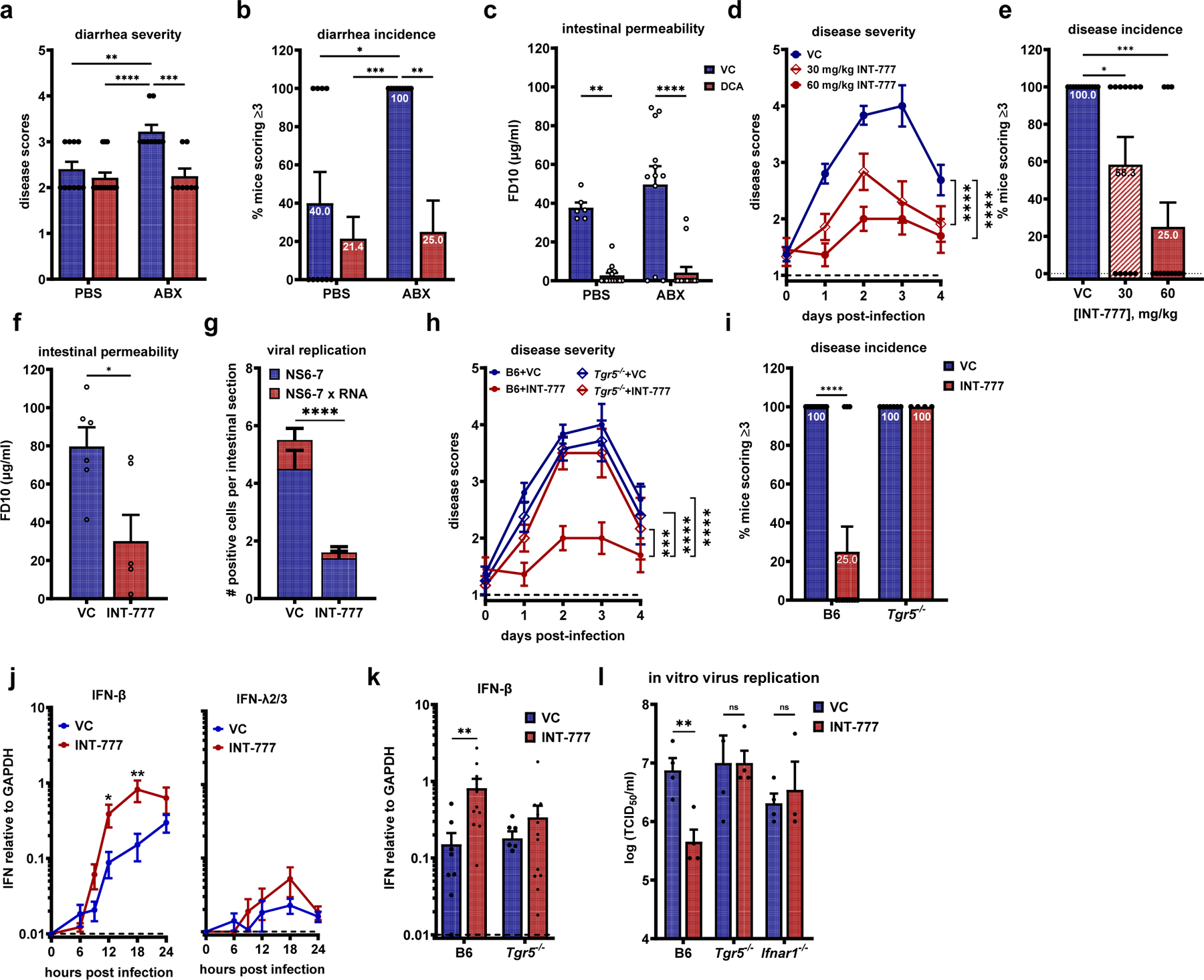
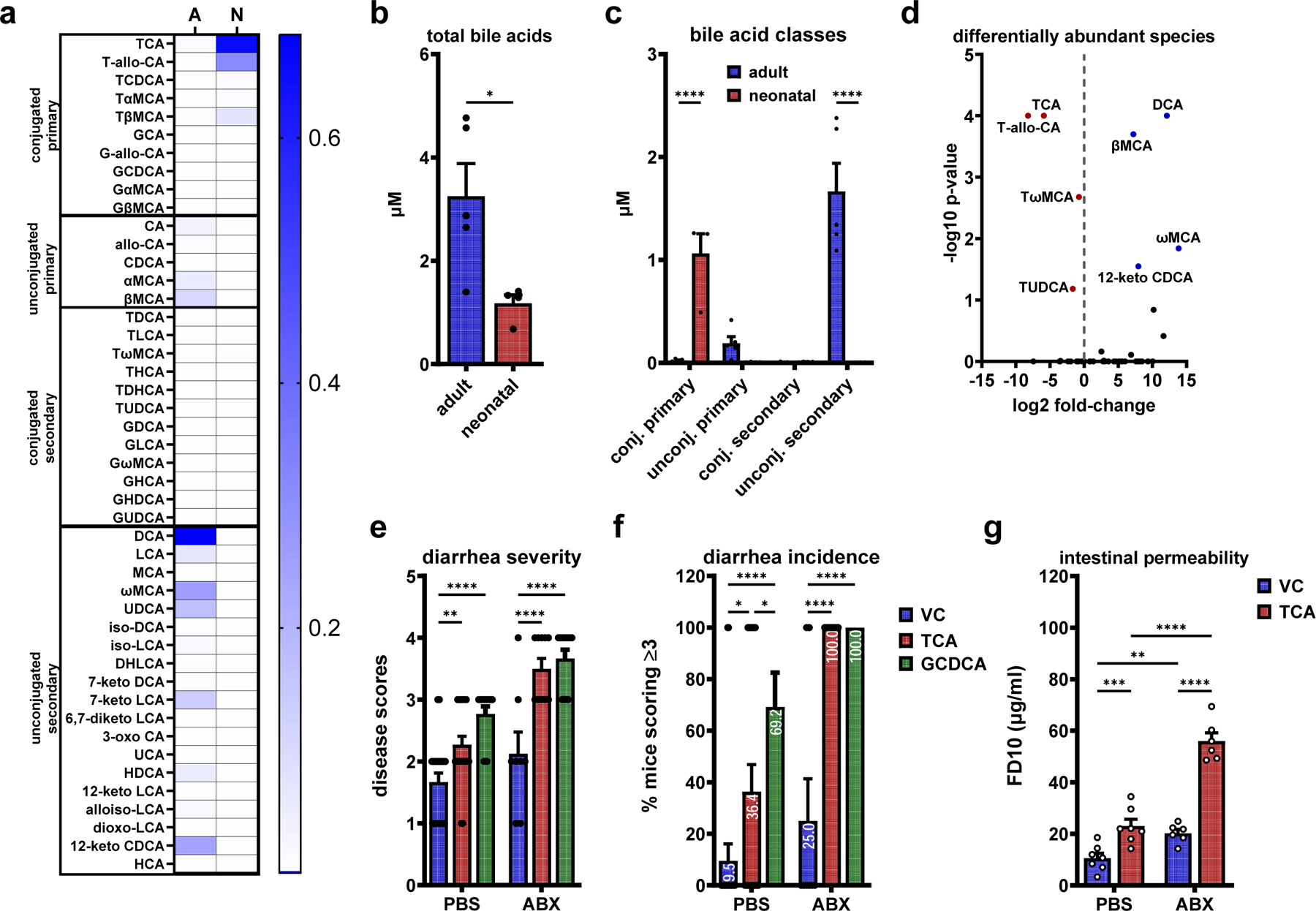
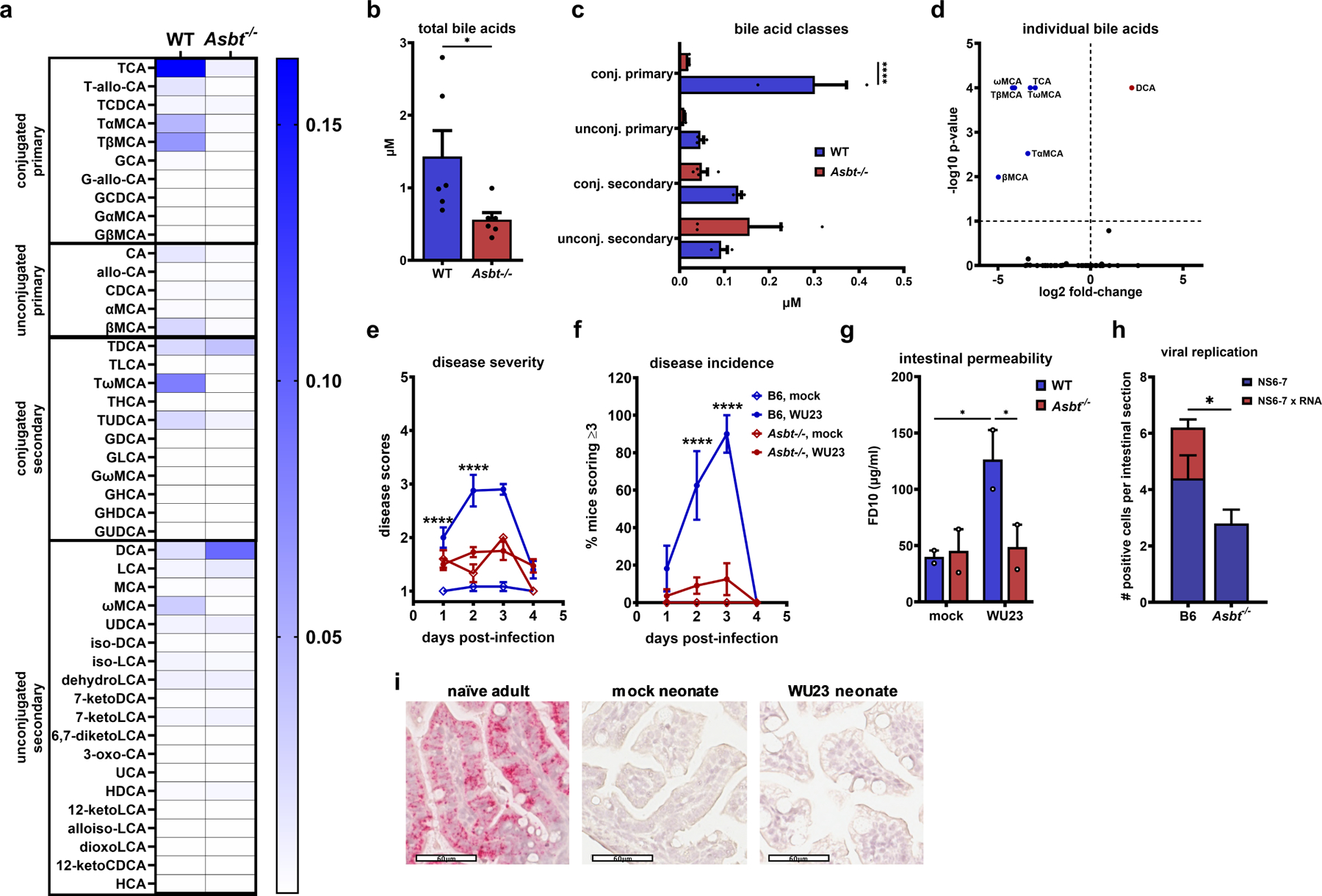
The pathogenic outcome of enteric virus infections is governed by a complex interplay between the virus, intestinal microbiota, and host immune factors, with metabolites serving as a key mediator. Noroviruses bind bile acid metabolites, which are produced by the host and then modified by commensal bacteria. Paradoxically, bile acids can have both proviral and antiviral roles during norovirus infections. Working in an infant mouse model of norovirus infection, we demonstrate that microbiota and their bile acid metabolites protect from norovirus diarrhea, whereas host bile acids promote disease. We also find that maternal bile acid metabolism determines the susceptibility of newborn mice to norovirus diarrhea during breastfeeding. Finally, targeting maternal and neonatal bile acid metabolism can protect newborn mice from norovirus disease. In summary, neonatal metabolic immaturity and breastmilk bile acids are central determinants of heightened newborn vulnerability to norovirus disease.
Keywords: ASBT; bile acids; breastmilk metabolites; enteromammary; gut-mammary; microbial metabolites; microbiota; neonatal infections; newborn infections; norovirus.
Copyright © 2024 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures


Update of
-
Metabolic immaturity of newborns and breast milk bile acid metabolites are the central determinants of heightened neonatal vulnerability to norovirus diarrhea.bioRxiv [Preprint]. 2024 May 1:2024.05.01.592031. doi: 10.1101/2024.05.01.592031. bioRxiv. 2024. Update in: Cell Host Microbe. 2024 Sep 11;32(9):1488-1501.e5. doi: 10.1016/j.chom.2024.08.003. PMID: 38746153 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
