

Nonstructural protein 4 of human norovirus self-assembles into various membrane-bridging multimers
- PMID: 39214299
- PMCID: PMC11439542
- DOI: 10.1016/j.jbc.2024.107724
Nonstructural protein 4 of human norovirus self-assembles into various membrane-bridging multimers
Abstract
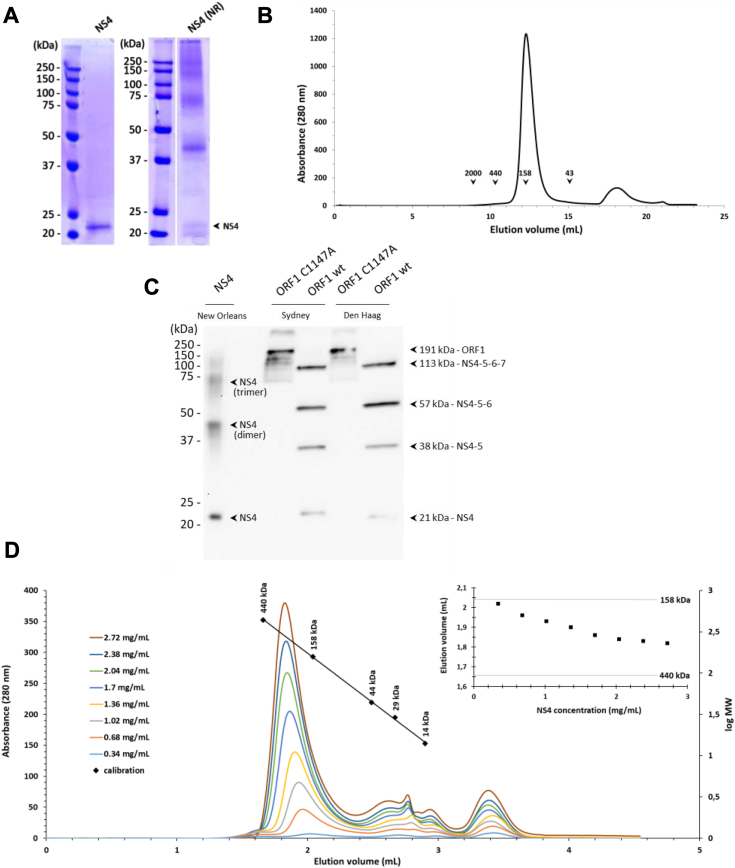
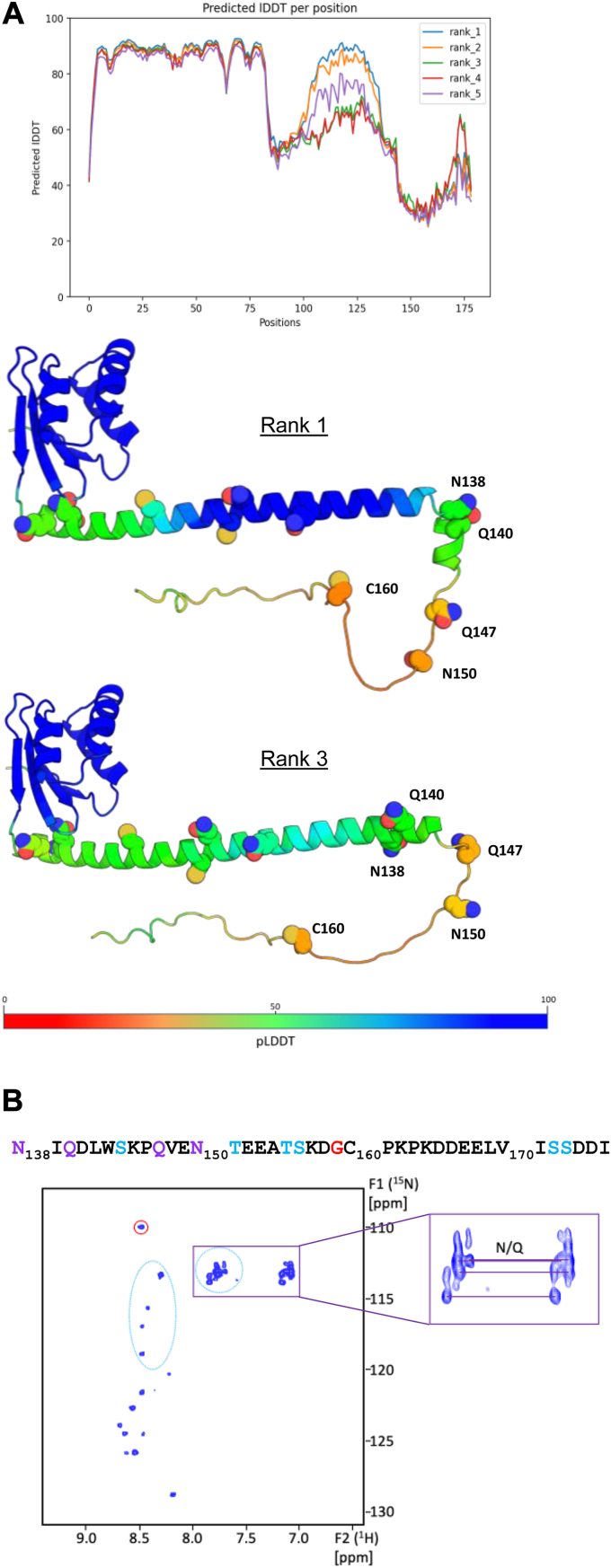
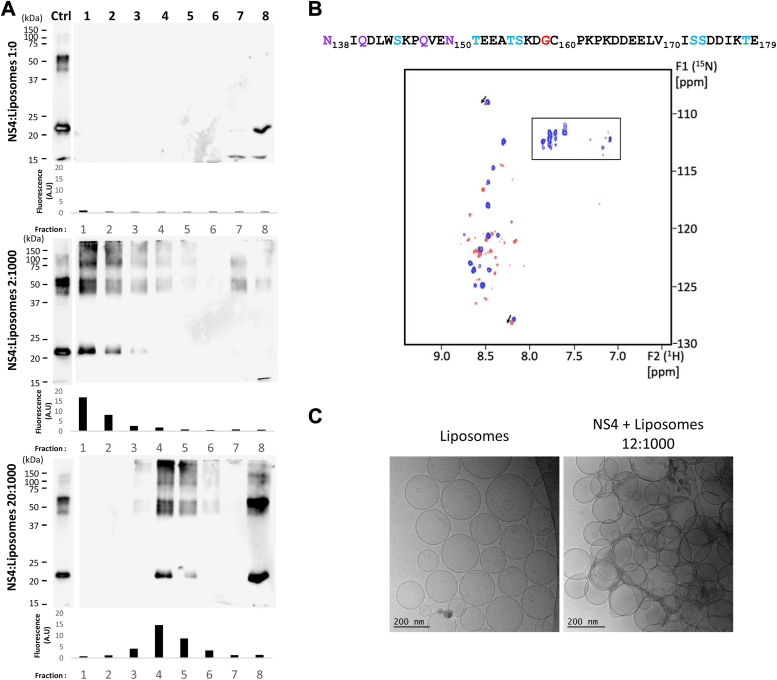
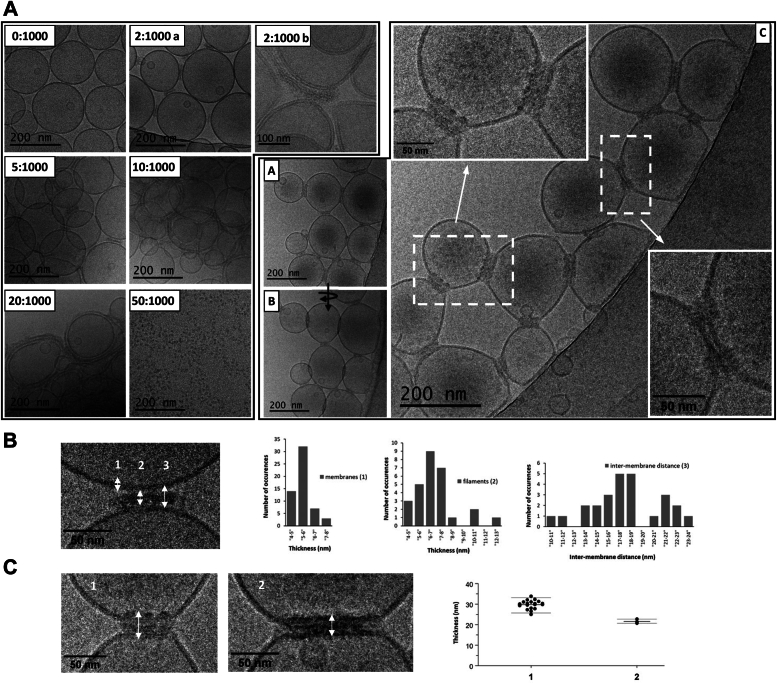
Single-stranded, positive-sense RNA ((+)RNA) viruses replicate their genomes in virus-induced intracellular membrane compartments. (+)RNA viruses dedicate a significant part of their small genomes (a few thousands to a few tens of thousands of bases) to the generation of these compartments by encoding membrane-interacting proteins and/or protein domains. Noroviruses are a very diverse genus of (+)RNA viruses including human and animal pathogens. Human noroviruses are the major cause of acute gastroenteritis worldwide, with genogroup II genotype 4 (GII.4) noroviruses accounting for the vast majority of infections. Three viral proteins encoded in the N terminus of the viral replication polyprotein direct intracellular membrane rearrangements associated with norovirus replication. Of these three, nonstructural protein 4 (NS4) seems to be the most important, although its exact functions in replication organelle formation are unknown. Here, we produce, purify, and characterize GII.4 NS4. AlphaFold modeling combined with experimental data refines and corrects our previous crude structural model of NS4. Using simple artificial liposomes, we report an extensive characterization of the membrane properties of NS4. We find that NS4 self-assembles and thereby bridges liposomes together. Cryo-EM, NMR, and membrane flotation show formation of several distinct NS4 assemblies, at least two of them bridging pairs of membranes together in different fashions. Noroviruses belong to (+)RNA viruses whose replication compartment is extruded from the target endomembrane and generates double-membrane vesicles. Our data establish that the 21-kDa GII.4 human norovirus NS4 can, in the absence of any other factor, recapitulate in tubo several features, including membrane apposition, that occur in such processes.
Keywords: cryo-EM; lipid–protein interaction; liposome; membrane; nonstructural protein; norovirus; plus-stranded RNA virus; protein assembly; viral protein; viral replication.
Copyright © 2024 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
