

Ion occupancy of the selectivity filter controls opening of a cytoplasmic gate in the K2P channel TALK-2
- PMID: 39215031
- PMCID: PMC11364775
- DOI: 10.1038/s41467-024-51812-w
Ion occupancy of the selectivity filter controls opening of a cytoplasmic gate in the K2P channel TALK-2
Abstract
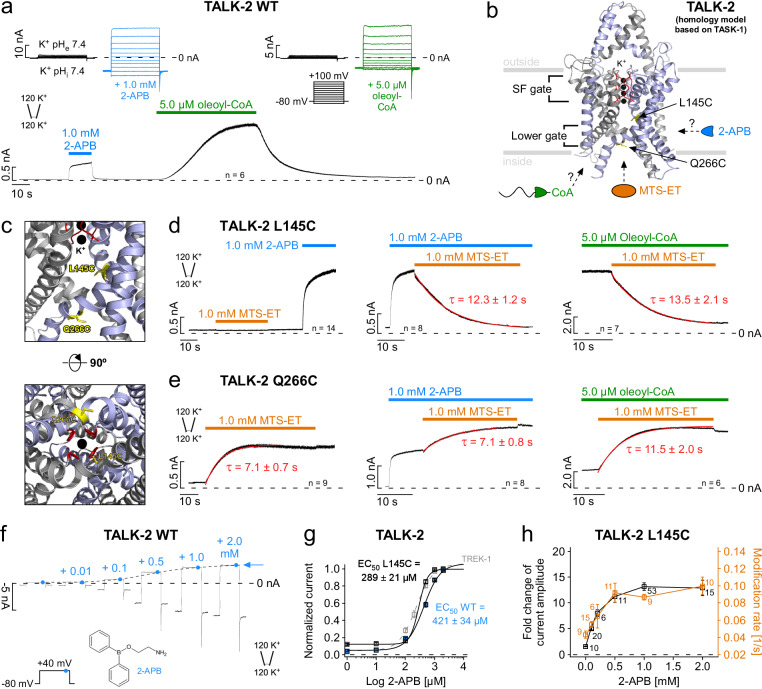
Two-pore domain K+ (K2P) channel activity was previously thought to be controlled primarily via a selectivity filter (SF) gate. However, recent crystal structures of TASK-1 and TASK-2 revealed a lower gate at the cytoplasmic pore entrance. Here, we report functional evidence of such a lower gate in the K2P channel K2P17.1 (TALK-2, TASK-4). We identified compounds (drugs and lipids) and mutations that opened the lower gate allowing the fast modification of pore cysteine residues. Surprisingly, stimuli that directly target the SF gate (i.e., pHe., Rb+ permeation, membrane depolarization) also opened the cytoplasmic gate. Reciprocally, opening of the lower gate reduced the electric work to open the SF via voltage driven ion binding. Therefore, it appears that the SF is so rigidly locked into the TALK-2 protein structure that changes in ion occupancy can pry open a distant lower gate and, vice versa, opening of the lower gate concurrently promote SF gate opening. This concept might extent to other K+ channels that contain two gates (e.g., voltage-gated K+ channels) for which such a positive gate coupling has been suggested, but so far not directly demonstrated.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures






References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
