

Human neural stem cells derived from fetal human brain communicate with each other and rescue ischemic neuronal cells through tunneling nanotubes
- PMID: 39217148
- PMCID: PMC11365985
- DOI: 10.1038/s41419-024-07005-w
Human neural stem cells derived from fetal human brain communicate with each other and rescue ischemic neuronal cells through tunneling nanotubes
Abstract
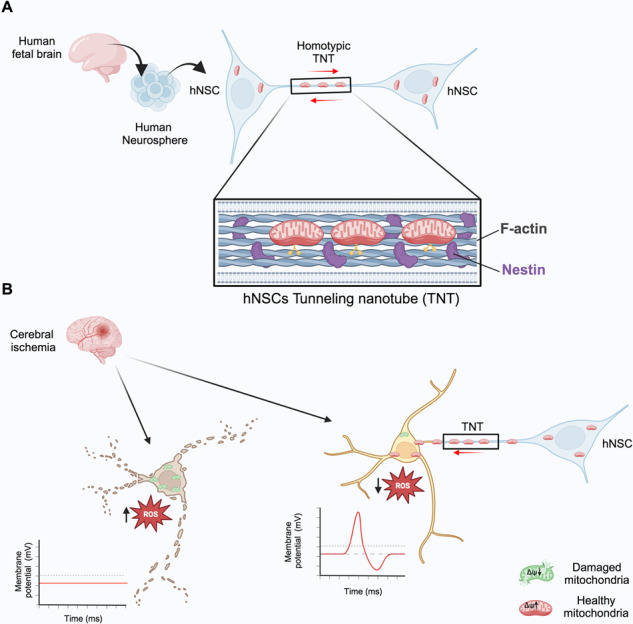
Pre-clinical trials have demonstrated the neuroprotective effects of transplanted human neural stem cells (hNSCs) during the post-ischemic phase. However, the exact neuroprotective mechanism remains unclear. Tunneling nanotubes (TNTs) are long plasma membrane bridges that physically connect distant cells, enabling the intercellular transfer of mitochondria and contributing to post-ischemic repair processes. Whether hNSCs communicate through TNTs and their role in post-ischemic neuroprotection remains unknown. In this study, non-immortalized hNSC lines derived from fetal human brain tissues were examined to explore these possibilities and assess the post-ischemic neuroprotection potential of these hNSCs. Using Tau-STED super-resolution confocal microscopy, live cell time-lapse fluorescence microscopy, electron microscopy, and direct or non-contact homotypic co-cultures, we demonstrated that hNSCs generate nestin-positive TNTs in both 3D neurospheres and 2D cultures, through which they transfer functional mitochondria. Co-culturing hNSCs with differentiated SH-SY5Y (dSH-SY5Y) revealed heterotypic TNTs allowing mitochondrial transfer from hNSCs to dSH-SY5Y. To investigate the role of heterotypic TNTs in post-ischemic neuroprotection, dSH-SY5Y were subjected to oxygen-glucose deprivation (OGD) followed by reoxygenation (OGD/R) with or without hNSCs in direct or non-contact co-cultures. Compared to normoxia, OGD/R dSH-SY5Y became apoptotic with impaired electrical activity. When OGD/R dSH-SY5Y were co-cultured in direct contact with hNSCs, heterotypic TNTs enabled the transfer of functional mitochondria from hNSCs to OGD/R dSH-SY5Y, rescuing them from apoptosis and restoring the bioelectrical profile toward normoxic dSH-SY5Y. This complete neuroprotection did not occur in the non-contact co-culture. In summary, our data reveal the presence of a functional TNTs network containing nestin within hNSCs, demonstrate the involvement of TNTs in post-ischemic neuroprotection mediated by hNSCs, and highlight the strong efficacy of our hNSC lines in post-ischemic neuroprotection. Human neural stem cells (hNSCs) communicate with each other and rescue ischemic neurons through nestin-positive tunneling nanotubes (TNTs). A Functional mitochondria are exchanged via TNTs between hNSCs. B hNSCs transfer functional mitochondria to ischemic neurons through TNTs, rescuing neurons from ischemia/reperfusion ROS-dependent apoptosis.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures








References
-
- Mendez I, Sanchez-Pernaute R, Cooper O, Viñuela A, Ferrari D, Björklund L. et al. Cell type analysis of functional fetal dopamine cell suspension transplants in the striatum and substantia nigra of patients with Parkinson’s disease. Brain. 2005;128(Part 7):1498–510. 10.1093/brain/awh510 - DOI - PMC - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
