

Perpetual step-like restructuring of hippocampal circuit dynamics
- PMID: 39217613
- PMCID: PMC11485410
- DOI: 10.1016/j.celrep.2024.114702
Perpetual step-like restructuring of hippocampal circuit dynamics
Abstract
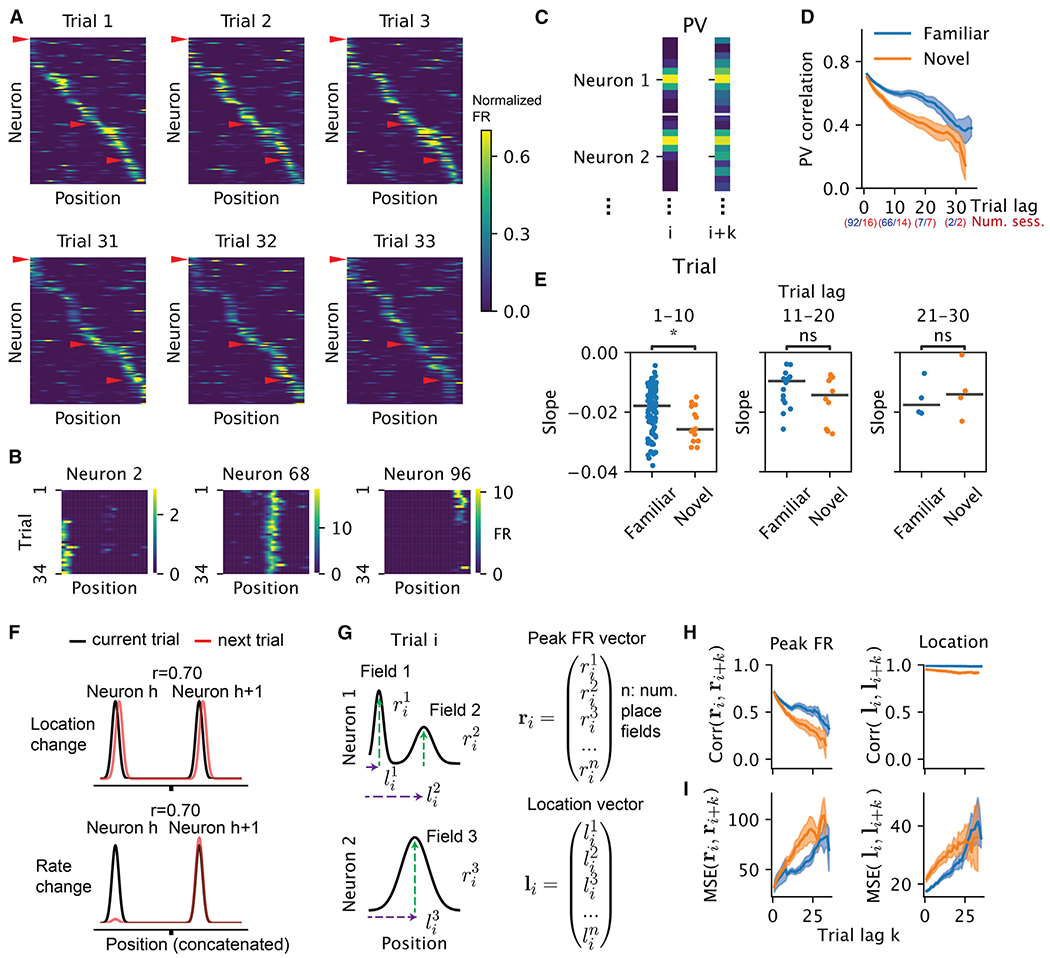
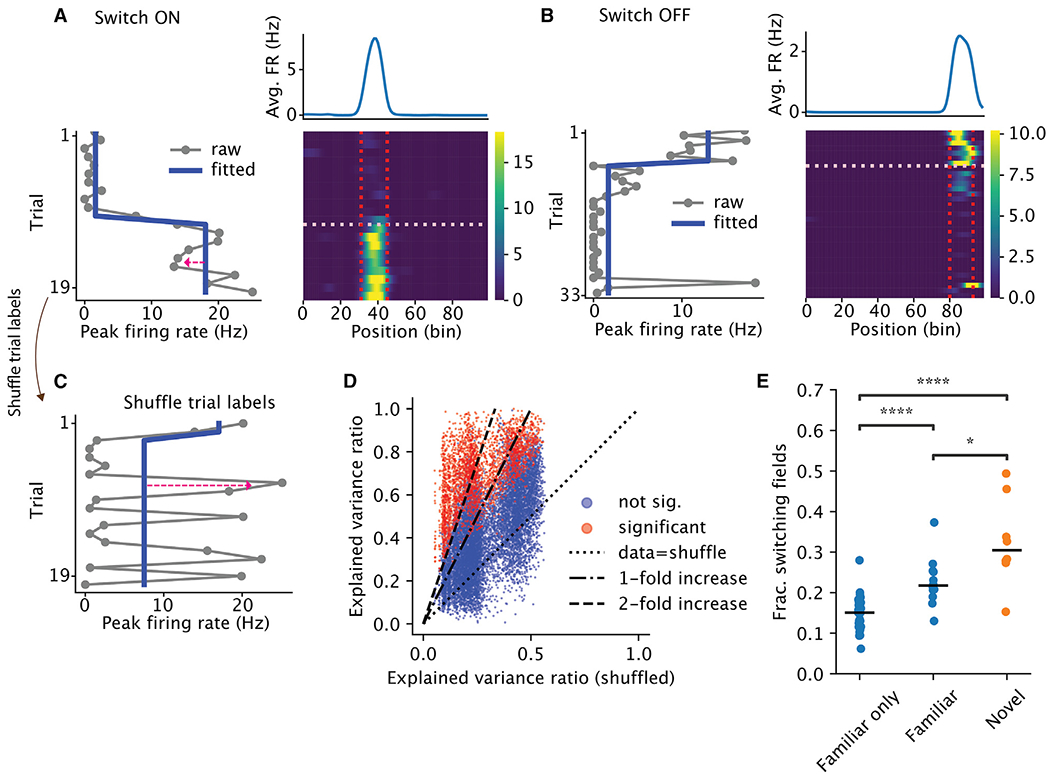
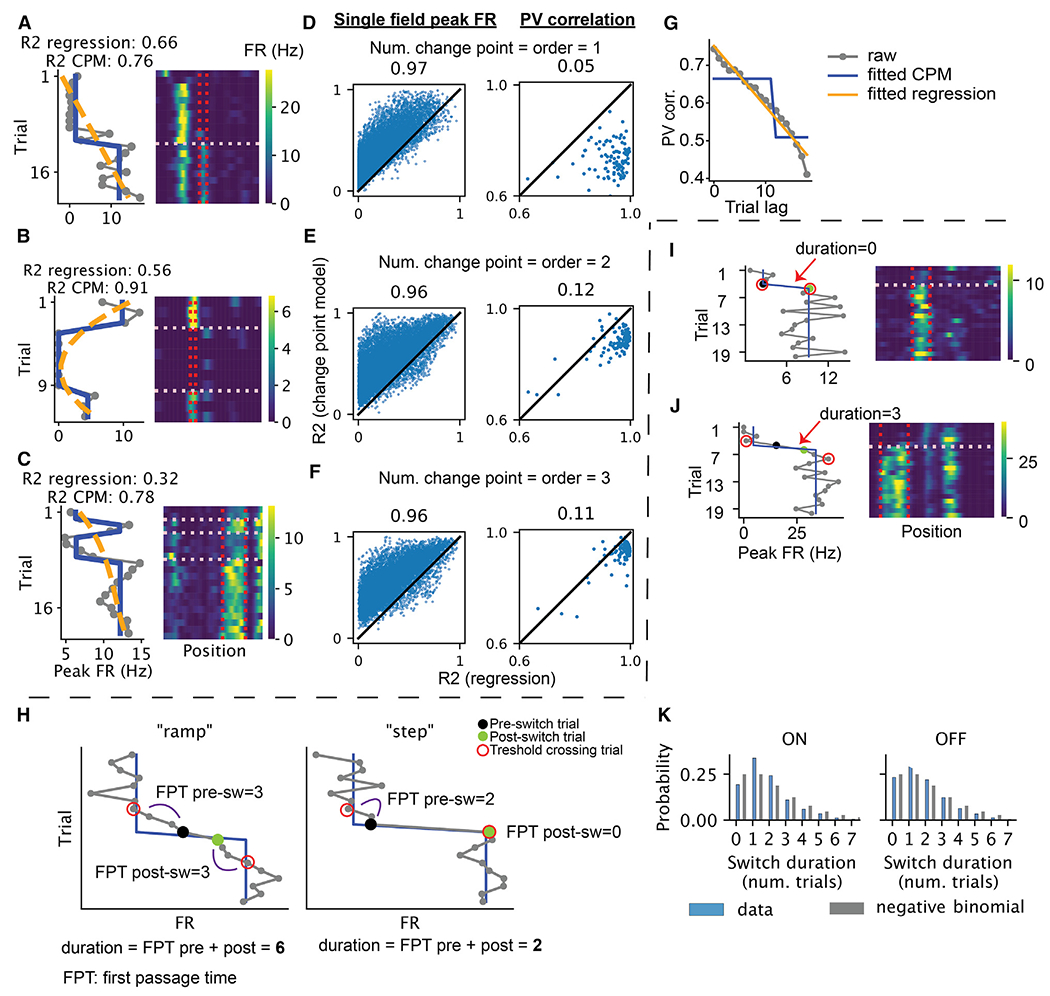
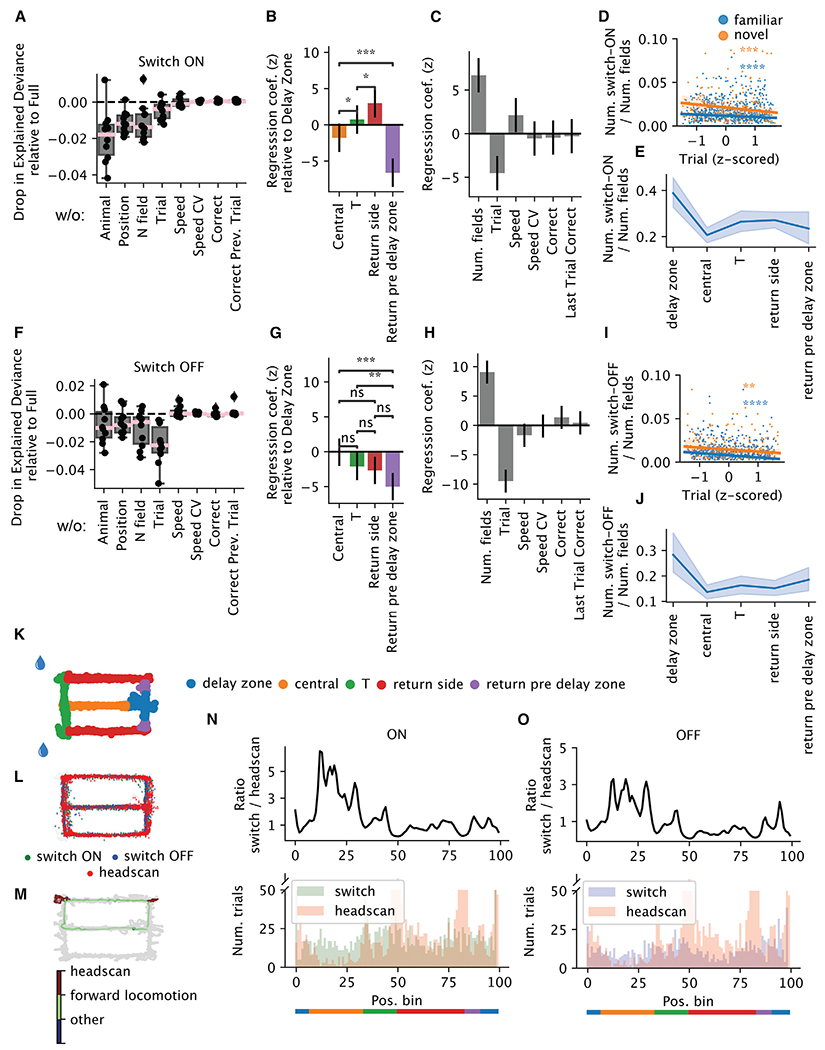
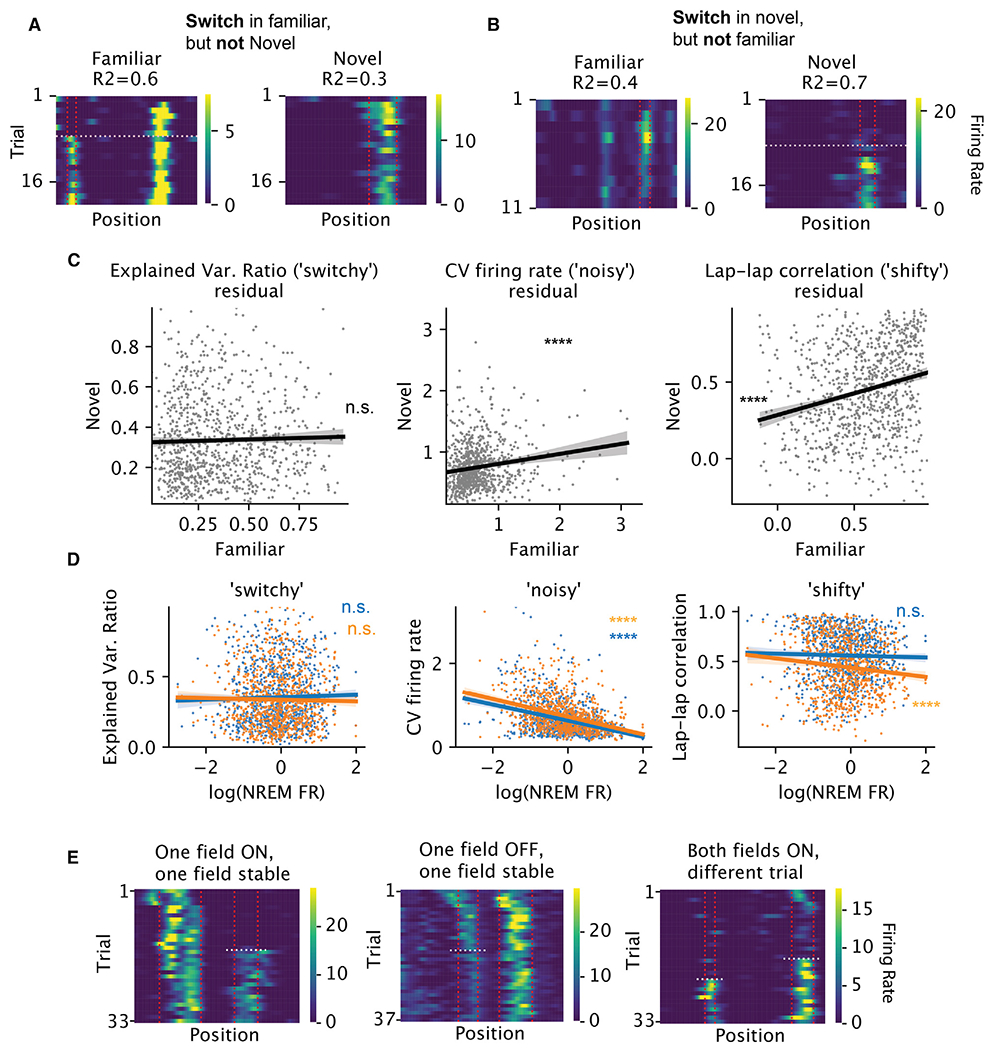
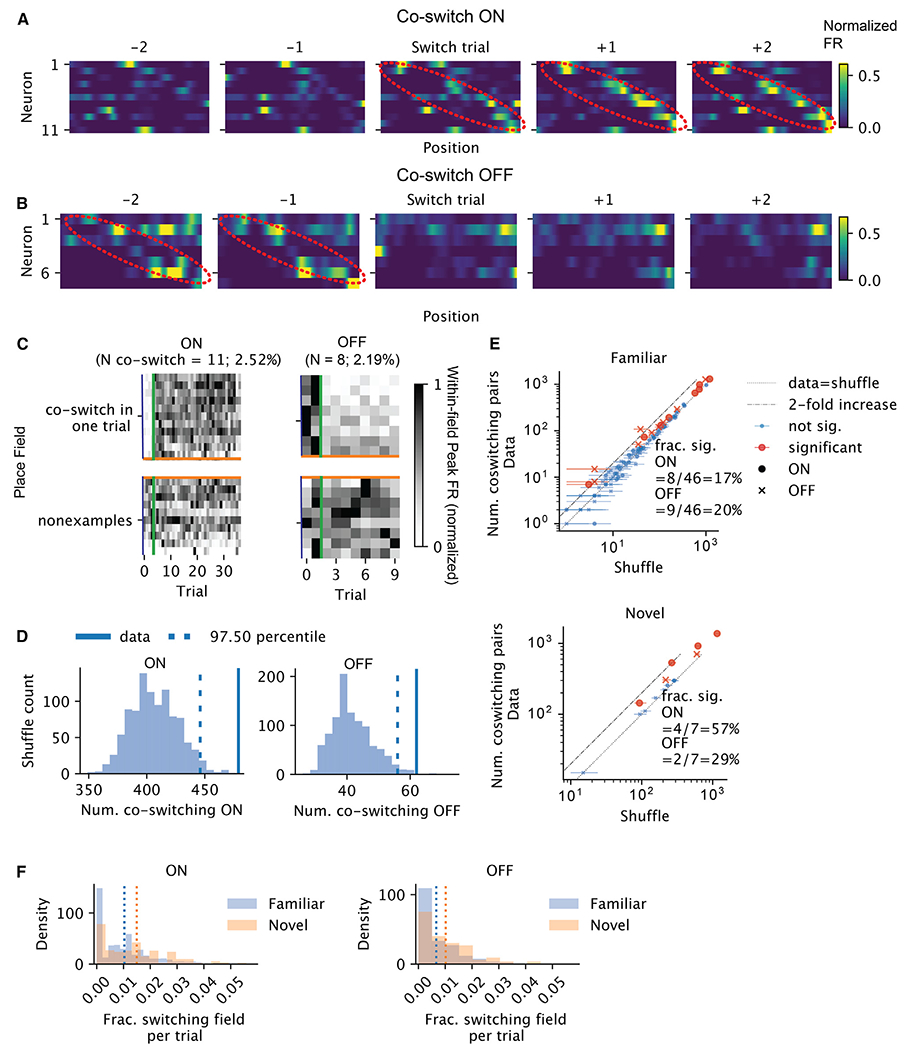
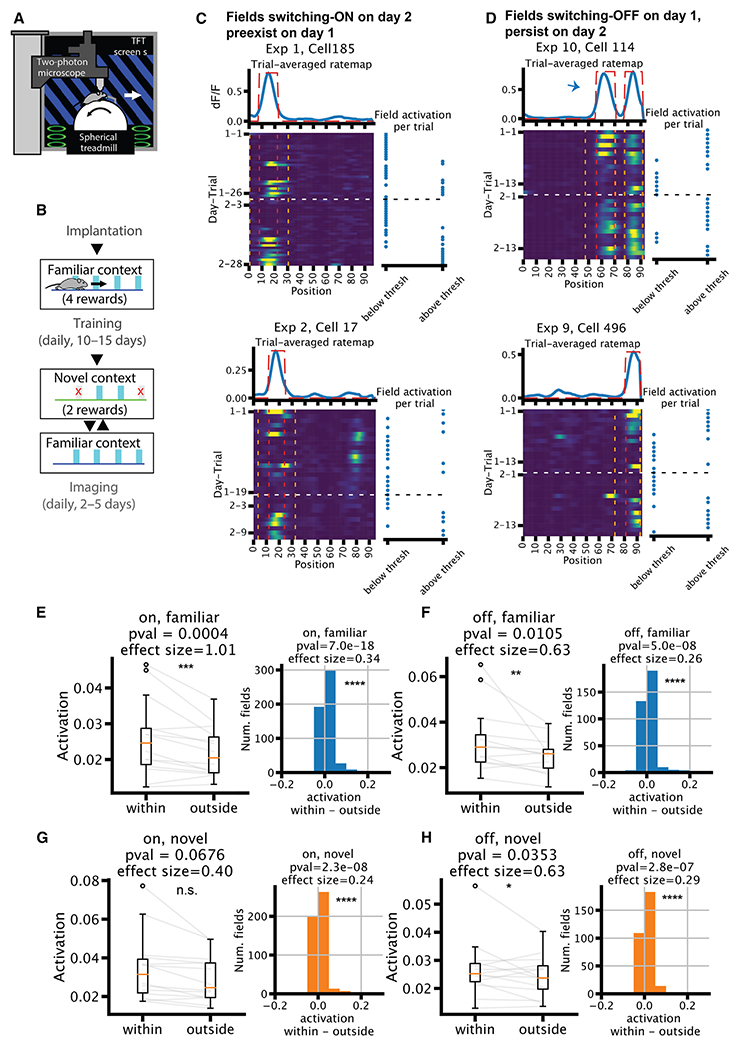
Representation of the environment by hippocampal populations is known to drift even within a familiar environment, which could reflect gradual changes in single-cell activity or result from averaging across discrete switches of single neurons. Disambiguating these possibilities is crucial, as they each imply distinct mechanisms. Leveraging change point detection and model comparison, we find that CA1 population vectors decorrelate gradually within a session. In contrast, individual neurons exhibit predominantly step-like emergence and disappearance of place fields or sustained changes in within-field firing. The changes are not restricted to particular parts of the maze or trials and do not require apparent behavioral changes. The same place fields emerge, disappear, and reappear across days, suggesting that the hippocampus reuses pre-existing assemblies, rather than forming new fields de novo. Our results suggest an internally driven perpetual step-like reorganization of the neuronal assemblies.
Keywords: CP: Neuroscience; change point detection; place fields; plasticity; pre-existing assemblies; quantal change; remapping; representational drift.
Copyright © 2024 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests G.B. is a member of the advisory board of Neuron.
Figures
Update of
-
Perpetual step-like restructuring of hippocampal circuit dynamics.bioRxiv [Preprint]. 2024 Apr 23:2024.04.22.590576. doi: 10.1101/2024.04.22.590576. bioRxiv. 2024. Update in: Cell Rep. 2024 Sep 24;43(9):114702. doi: 10.1016/j.celrep.2024.114702. PMID: 38712105 Free PMC article. Updated. Preprint.
References
-
- O’Keefe J, and Nadel L (1978). The Hippocampus as a Cognitive Map (Clarendon Press; ).
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
