

Fungal Peptidomelanin: A Novel Biopolymer for the Chelation of Heavy Metals
- PMID: 39220543
- PMCID: PMC11359623
- DOI: 10.1021/acsomega.4c03704
Fungal Peptidomelanin: A Novel Biopolymer for the Chelation of Heavy Metals
Abstract
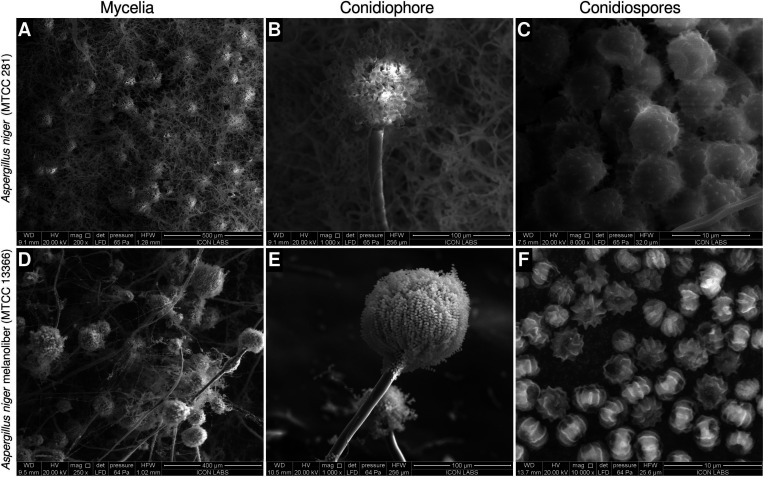
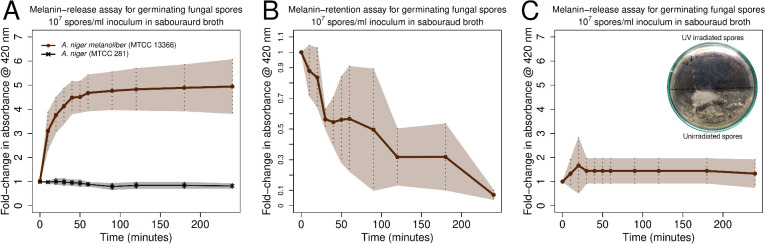
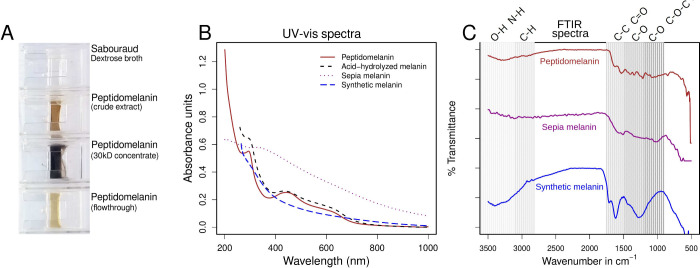
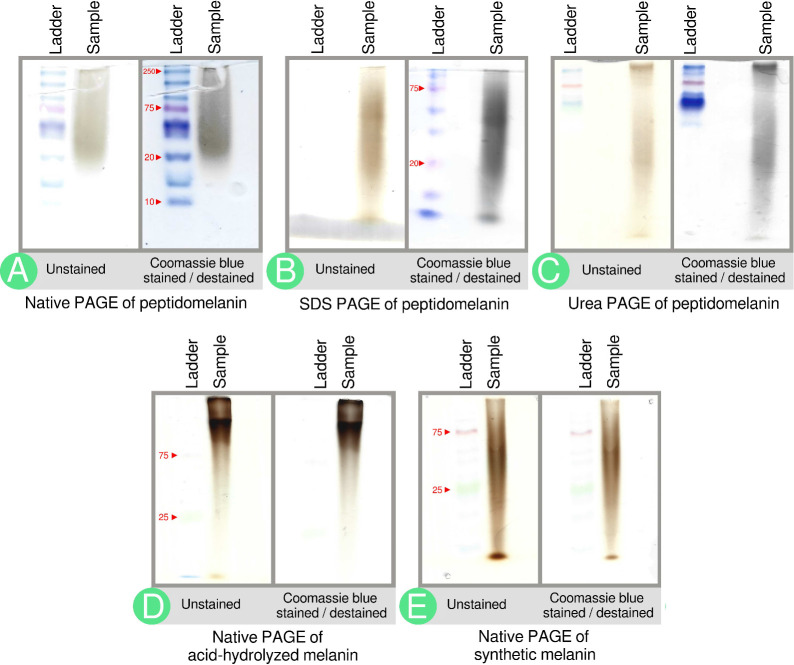
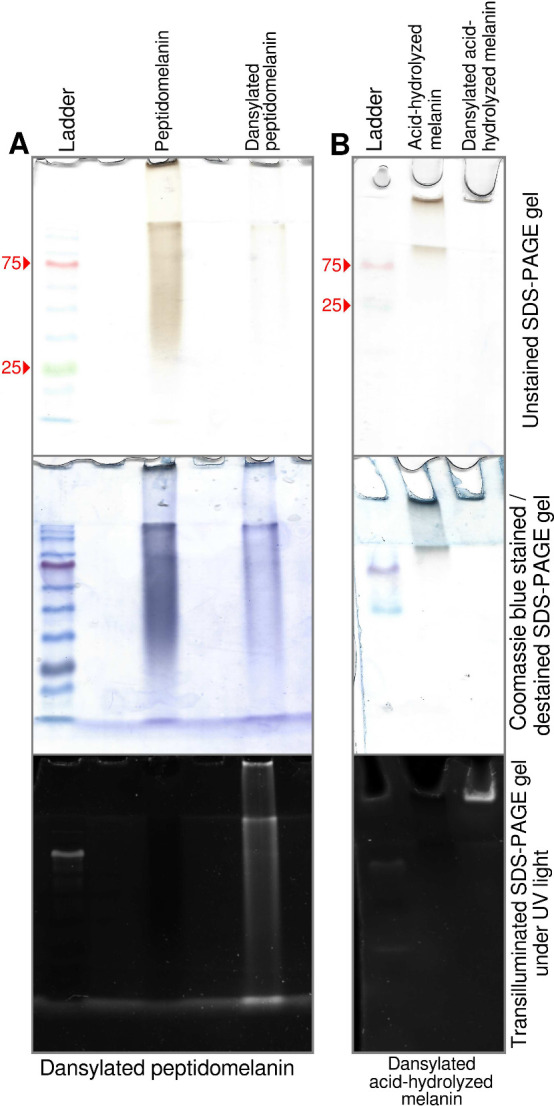
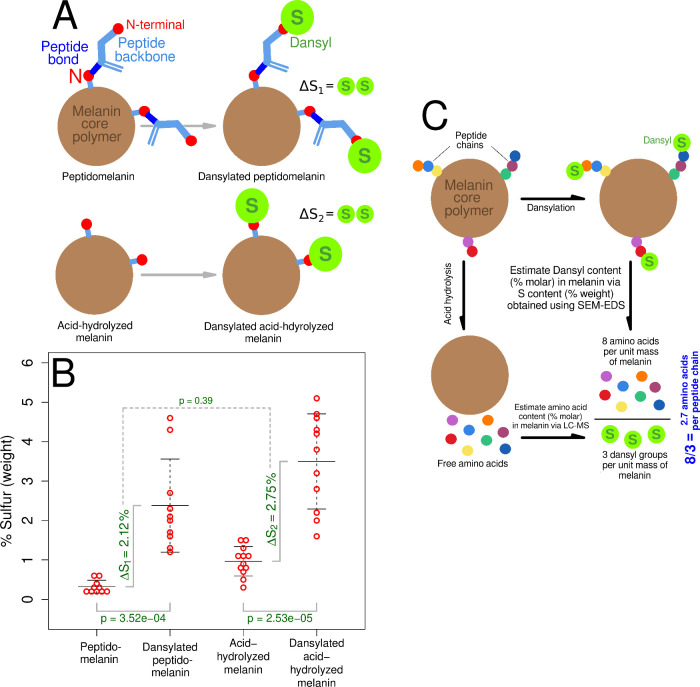
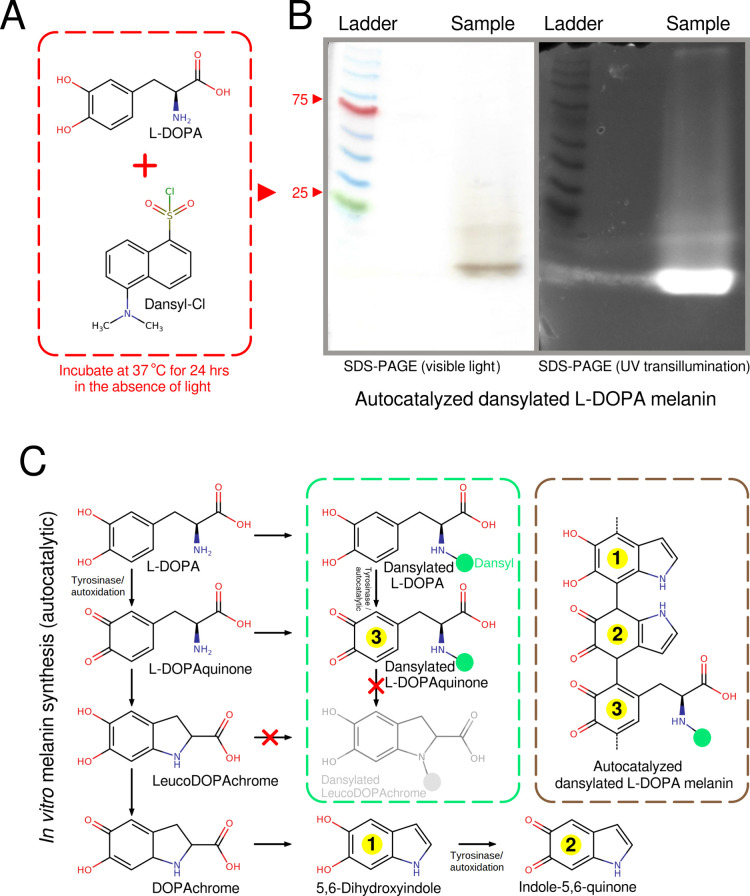
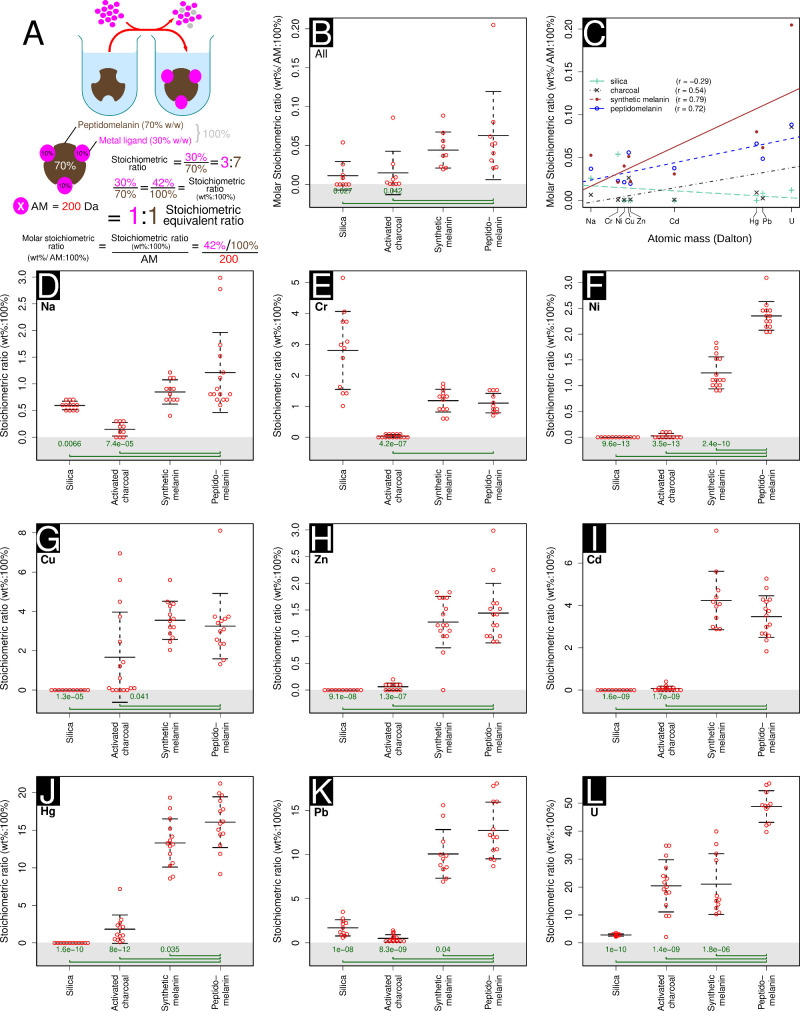
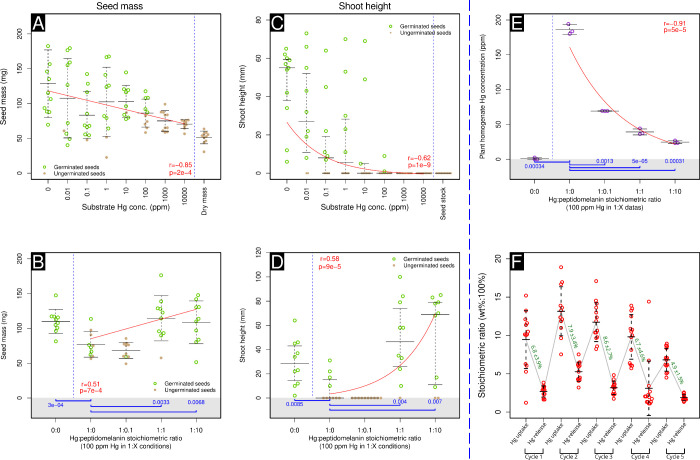
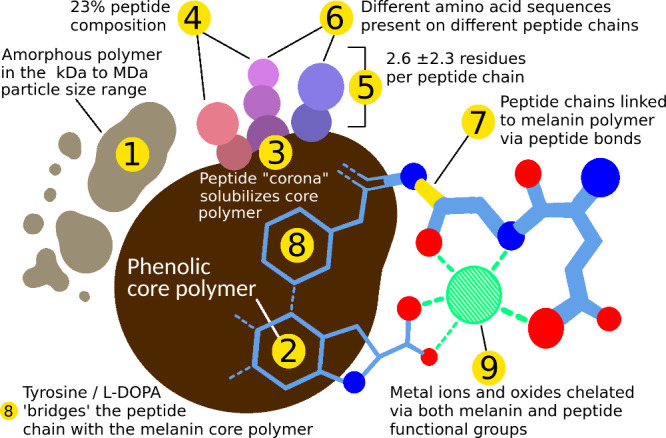
Melanin is an amorphous, highly heterogeneous polymer found across all kingdoms of life. Although the properties of melanin can greatly vary, most forms are insoluble and strongly absorb light, appearing dark brown to black. Here, we describe a water-soluble form of melanin (peptidomelanin) secreted by the spores of Aspergillus niger (strain: melanoliber) during germination. Peptidomelanin is composed of an insoluble L-DOPA core polymer that is solubilized via short, copolymerized heterogeneous peptide chains forming a "corona" with a mean amino acid length of 2.6 ± 2.3. Based on in vitro experiments, we propose a biochemical copolymerization mechanism involving the hydroxylation of tyrosynylated peptides. Peptidomelanin is capable of chelating heavy metals such as lead, mercury, and uranium (as uranyl) in large quantities. Preliminary data indicates that peptidomelanin may have applications for the remediation of heavy metals in situ, including in agricultural settings.
© 2024 The Authors. Published by American Chemical Society.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
References
-
- Andersen N.; et al. Structure, chemistry, and biosynthesis of the melanins. Fortschr. Chem. Org. Naturst. 1974, 521–582. - PubMed
LinkOut - more resources
Full Text Sources