

DMDA-PatA mediates RNA sequence-selective translation repression by anchoring eIF4A and DDX3 to GNG motifs
- PMID: 39223140
- PMCID: PMC11369270
- DOI: 10.1038/s41467-024-51635-9
DMDA-PatA mediates RNA sequence-selective translation repression by anchoring eIF4A and DDX3 to GNG motifs
Abstract
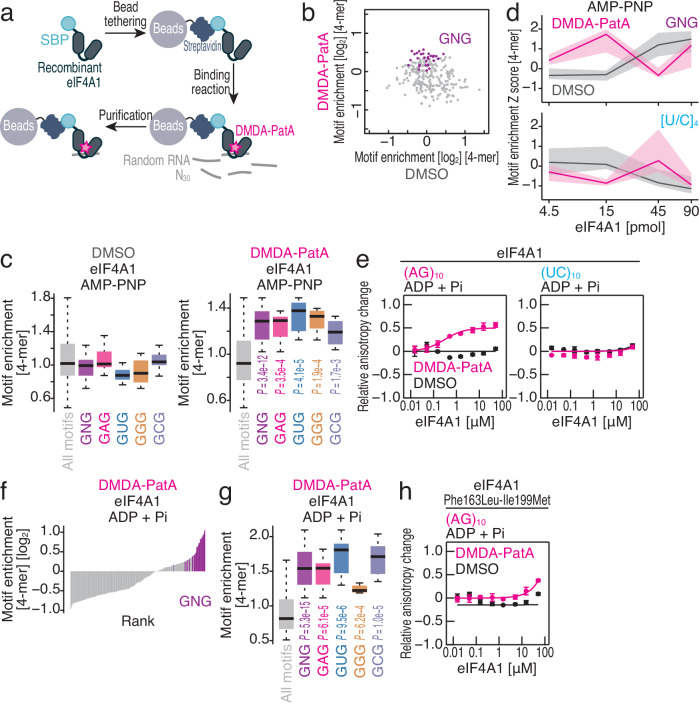
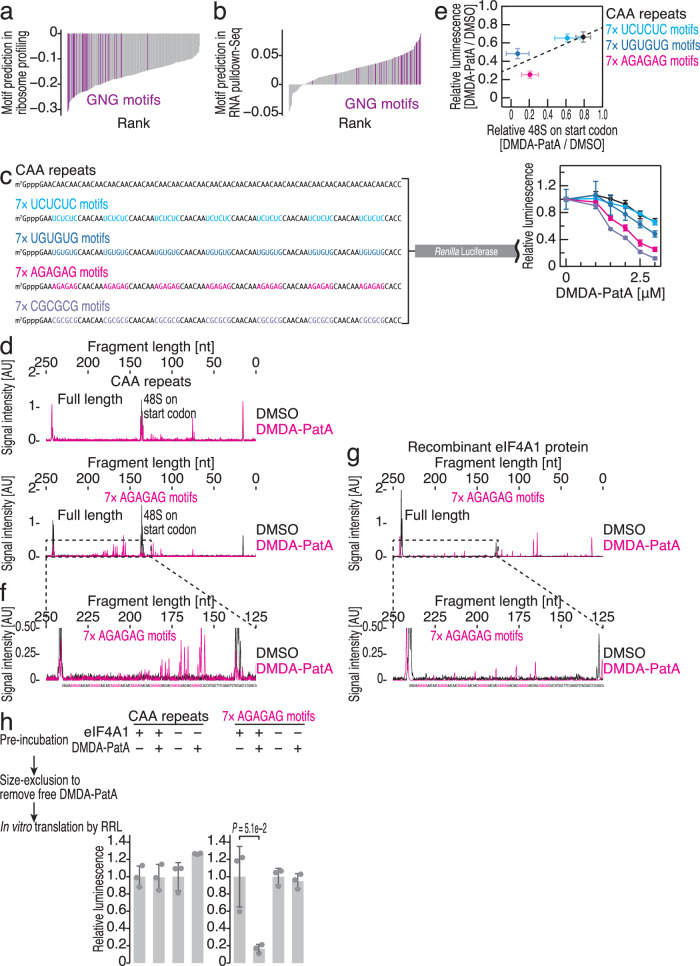
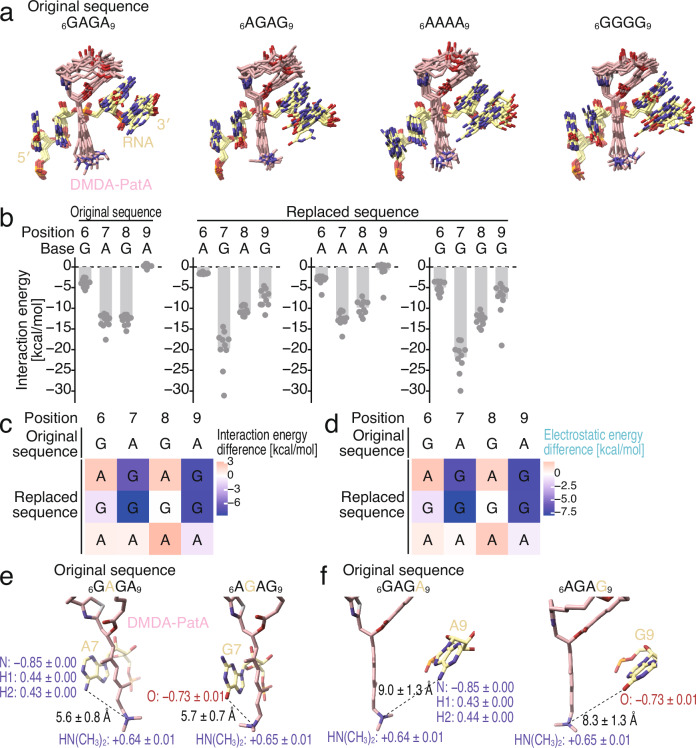
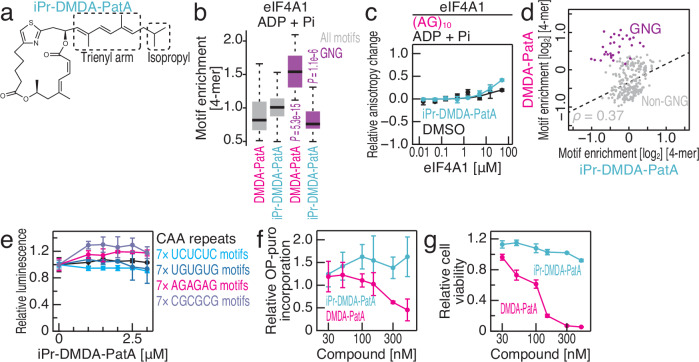
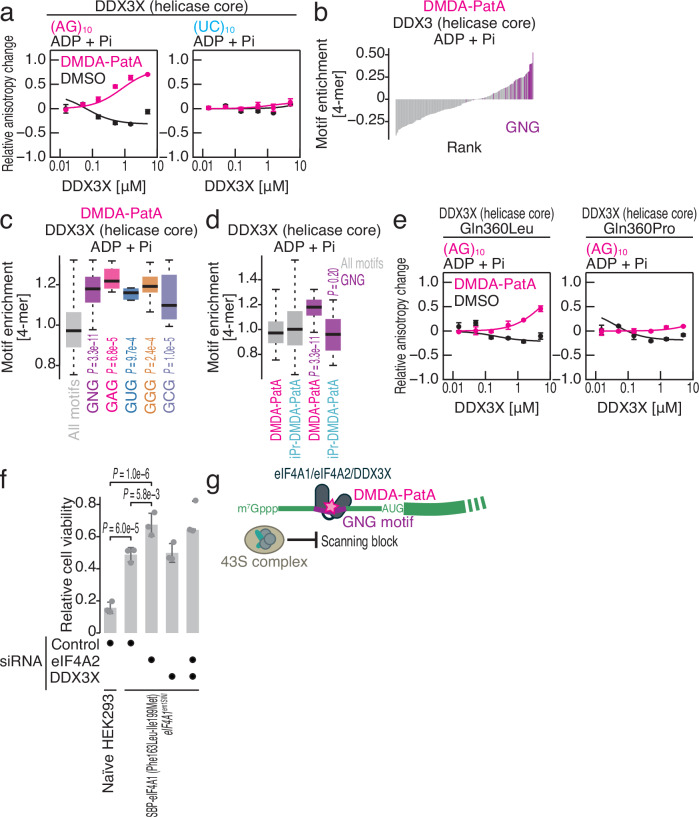
Small-molecule compounds that elicit mRNA-selective translation repression have attracted interest due to their potential for expansion of druggable space. However, only a limited number of examples have been reported to date. Here, we show that desmethyl desamino pateamine A (DMDA-PatA) represses translation in an mRNA-selective manner by clamping eIF4A, a DEAD-box RNA-binding protein, onto GNG motifs. By systematically comparing multiple eIF4A inhibitors by ribosome profiling, we found that DMDA-PatA has unique mRNA selectivity for translation repression. Unbiased Bind-n-Seq reveals that DMDA-PatA-targeted eIF4A exhibits a preference for GNG motifs in an ATP-independent manner. This unusual RNA binding sterically hinders scanning by 40S ribosomes. A combination of classical molecular dynamics simulations and quantum chemical calculations, and the subsequent development of an inactive DMDA-PatA derivative reveals that the positive charge of the tertiary amine on the trienyl arm induces G selectivity. Moreover, we identified that DDX3, another DEAD-box protein, is an alternative DMDA-PatA target with the same effects on eIF4A. Our results provide an example of the sequence-selective anchoring of RNA-binding proteins and the mRNA-selective inhibition of protein synthesis by small-molecule compounds.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures

References
MeSH terms
Substances
Grants and funding
- Incentive Research Projects/MEXT | RIKEN
- JP23gm1410001/Japan Agency for Medical Research and Development (AMED)
- JP23H00095/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP23H04268/Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- JP18H05503/Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- S10 OD018174/OD/NIH HHS/United States
- R01 GM052964/GM/NIGMS NIH HHS/United States
- JP21H05281/Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- Pioneering Projects/MEXT | RIKEN
- JP23K05648/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP19H05640/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP21H05734/Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- R37 GM052964/GM/NIGMS NIH HHS/United States
- JP23H02415/MEXT | Japan Society for the Promotion of Science (JSPS)
- JP24H02307/Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- JP20H05784/Ministry of Education, Culture, Sports, Science and Technology (MEXT)
- R29 GM052964/GM/NIGMS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
