

Epigenetic therapy potentiates transposable element transcription to create tumor-enriched antigens in glioblastoma cells
- PMID: 39223316
- PMCID: PMC12180585
- DOI: 10.1038/s41588-024-01880-x
Epigenetic therapy potentiates transposable element transcription to create tumor-enriched antigens in glioblastoma cells
Abstract
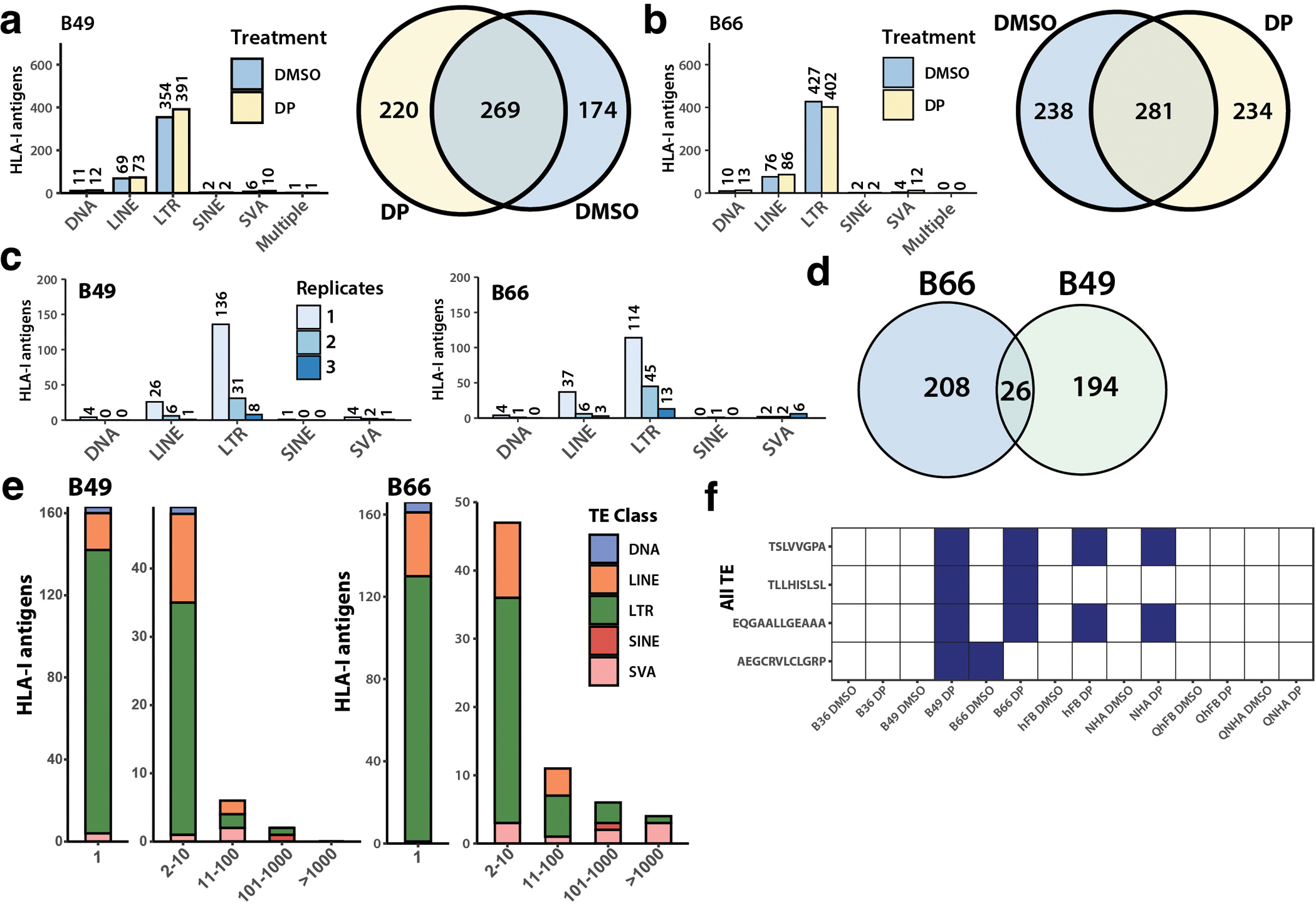
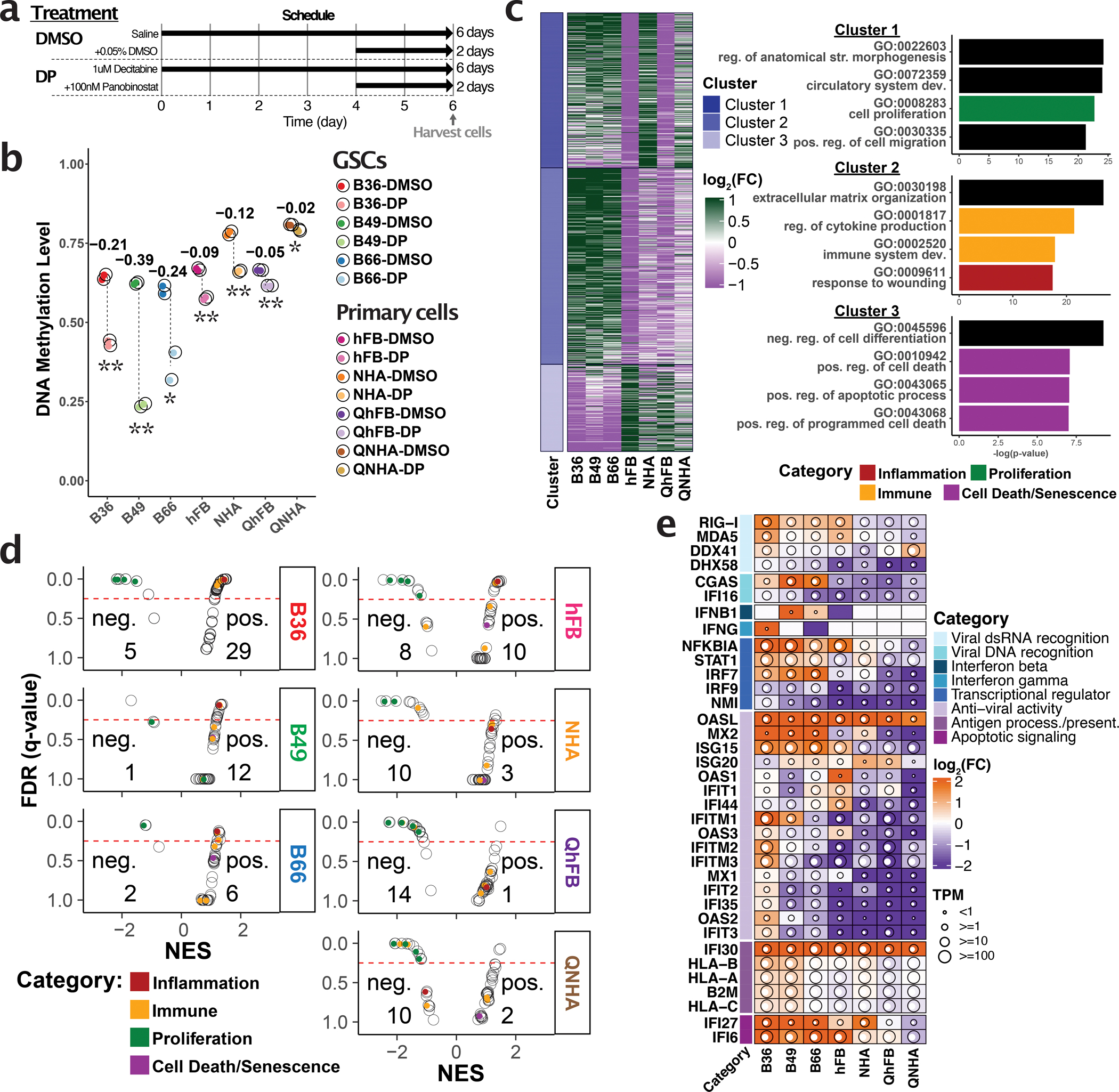
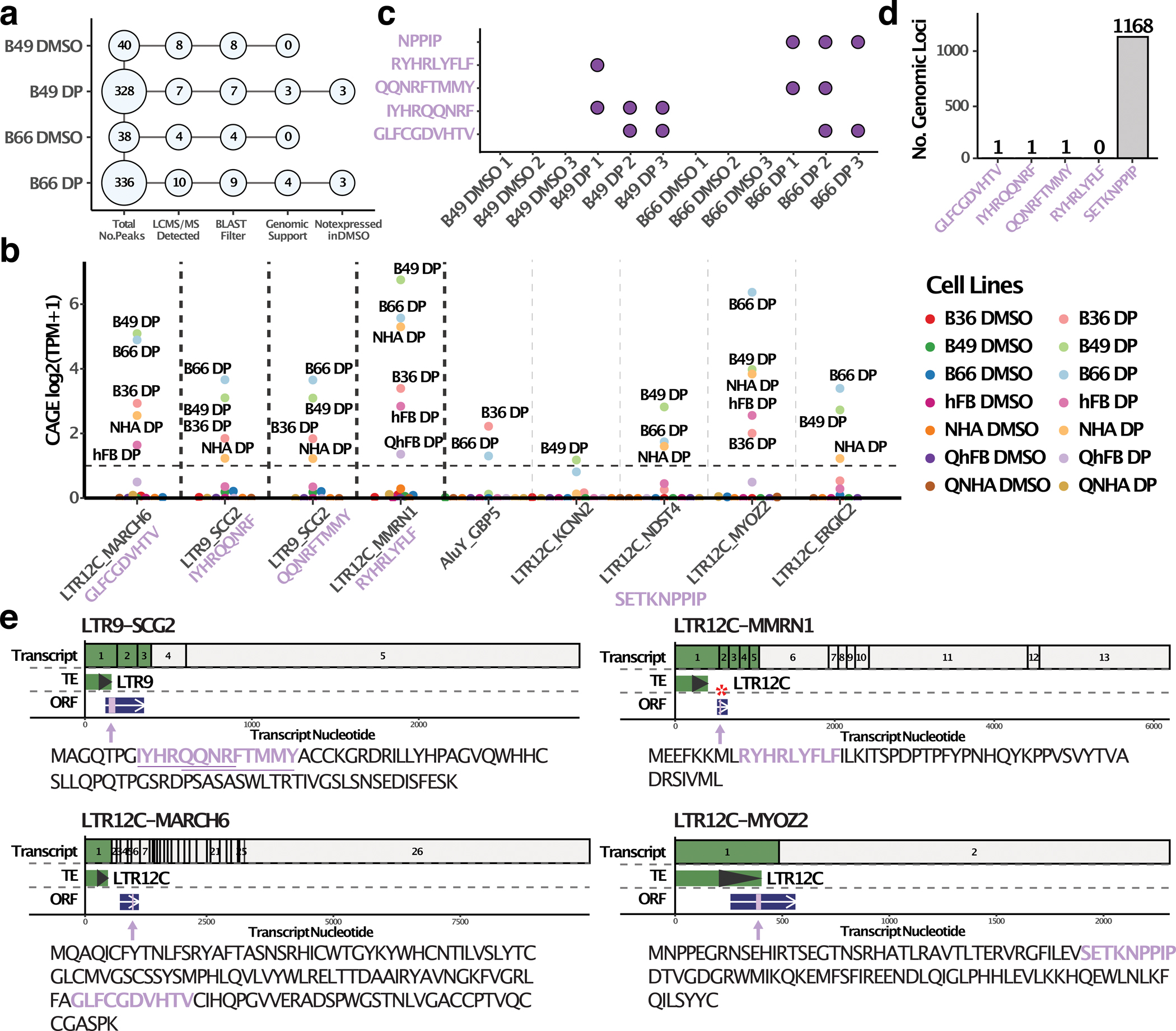
Inhibiting epigenetic modulators can transcriptionally reactivate transposable elements (TEs). These TE transcripts often generate unique peptides that can serve as immunogenic antigens for immunotherapy. Here, we ask whether TEs activated by epigenetic therapy could appreciably increase the antigen repertoire in glioblastoma, an aggressive brain cancer with low mutation and neoantigen burden. We treated patient-derived primary glioblastoma stem cell lines, an astrocyte cell line and primary fibroblast cell lines with epigenetic drugs, and identified treatment-induced, TE-derived transcripts that are preferentially expressed in cancer cells. We verified that these transcripts could produce human leukocyte antigen class I-presented antigens using liquid chromatography with tandem mass spectrometry pulldown experiments. Importantly, many TEs were also transcribed, even in proliferating nontumor cell lines, after epigenetic therapy, which suggests that targeted strategies like CRISPR-mediated activation could minimize potential side effects of activating unwanted genomic regions. The results highlight both the need for caution and the promise of future translational efforts in harnessing treatment-induced TE-derived antigens for targeted immunotherapy.
© 2024. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing Interests Statement
A.H.K. is a consultant for Monteris Medical. All other authors declare no competing interests.
Figures








References
-
- Gong J, Chehrazi-Raffle A, Reddi S & Salgia R Development of PD-1 and PD-L1 inhibitors as a form of cancer immunotherapy: A comprehensive review of registration trials and future considerations. Journal for ImmunoTherapy of Cancer vol. 6 Preprint at 10.1186/s40425-018-0316-z (2018). - DOI - PMC - PubMed
Methods-only References
-
- Liao Y, Smyth GK & Shi W FeatureCounts: An efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930 (2014). - PubMed
MeSH terms
Substances
Grants and funding
- U01 HG009391/HG/NHGRI NIH HHS/United States
- R01 HG007175/HG/NHGRI NIH HHS/United States
- T32 GM007200/GM/NIGMS NIH HHS/United States
- R01 NS128470/NS/NINDS NIH HHS/United States
- U01HG009391/U.S. Department of Health & Human Services | NIH | National Human Genome Research Institute (NHGRI)
- P30 CA091842/CA/NCI NIH HHS/United States
- P30 DK020579/DK/NIDDK NIH HHS/United States
- UL1 TR002345/TR/NCATS NIH HHS/United States
- 5R01HG007175/U.S. Department of Health & Human Services | NIH | National Human Genome Research Institute (NHGRI)
- RSG-14-049-01-DMC/American Cancer Society (American Cancer Society, Inc.)
- P60 DK020579/DK/NIDDK NIH HHS/United States
- U24ES026699/U.S. Department of Health & Human Services | NIH | National Institute of Environmental Health Sciences (NIEHS)
- T32 GM007067/GM/NIGMS NIH HHS/United States
- U24 ES026699/ES/NIEHS NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
