

LAMTOR1 decreased exosomal PD-L1 to enhance immunotherapy efficacy in non-small cell lung cancer
- PMID: 39223601
- PMCID: PMC11367890
- DOI: 10.1186/s12943-024-02099-4
LAMTOR1 decreased exosomal PD-L1 to enhance immunotherapy efficacy in non-small cell lung cancer
Abstract
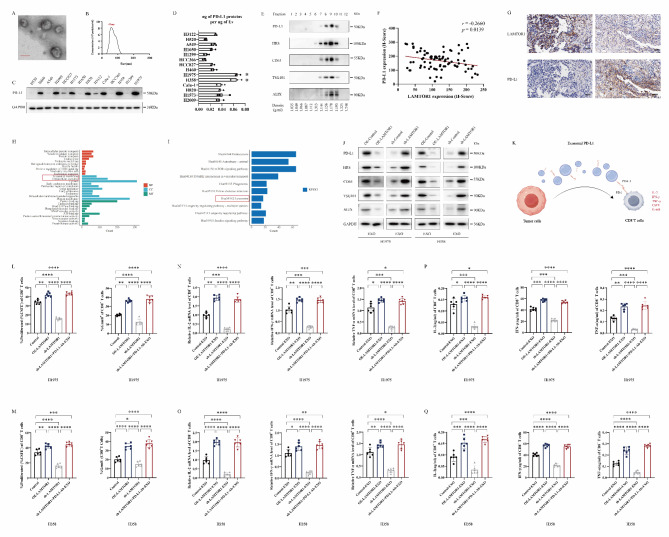
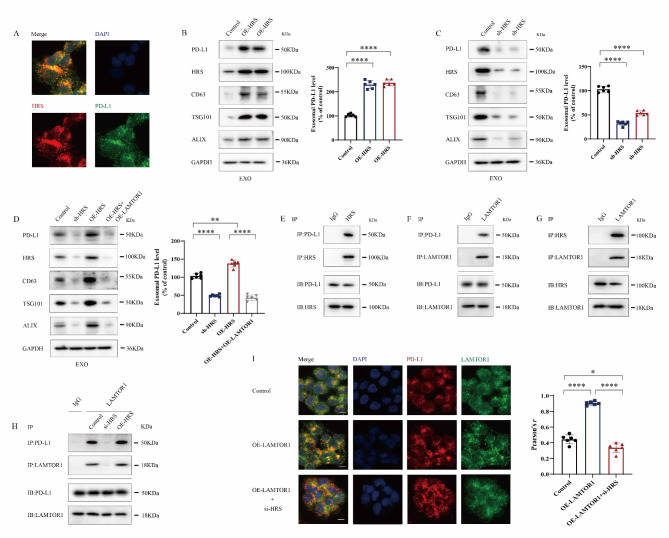
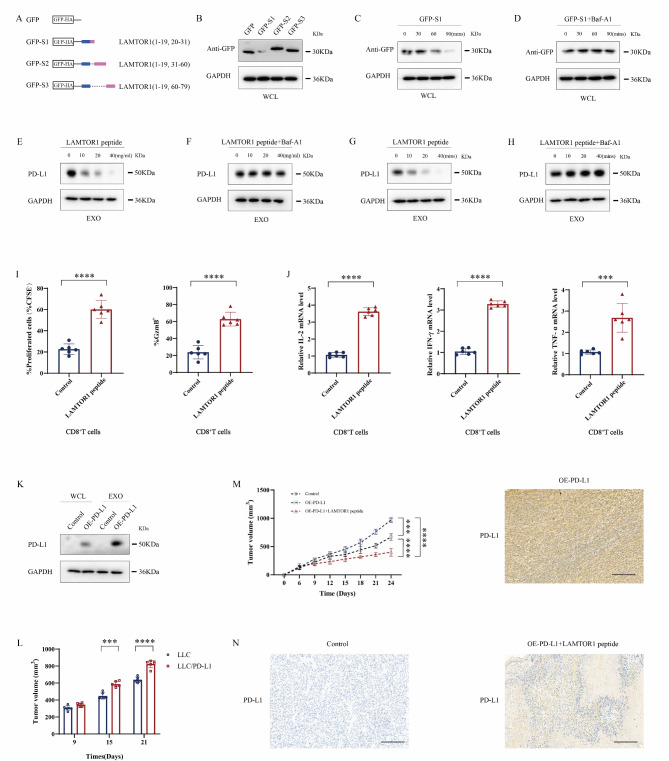
Great progress has been made in utilizing immune checkpoint blockade (ICB) for the treatment of non-small-cell lung cancer (NSCLC). Therapies targeting programmed cell death protein 1 (PD-1) and its ligand PD-L1, expressed on tumor cells, have demonstrated potential in improving patient survival rates. An unresolved issue involves immune suppression induced by exosomal PD-L1 within the tumor microenvironment (TME), particularly regarding CD8+ T cells. Our study unveiled the crucial involvement of LAMTOR1 in suppressing the exosomes of PD-L1 and promoting CD8+ T cell infiltration in NSCLC. Through its interaction with HRS, LAMTOR1 facilitates PD-L1 lysosomal degradation, thereby reducing exosomal PD-L1 release. Notably, the ability of LAMTOR1 to promote PD-L1 lysosomal degradation relies on a specific ubiquitination site and an HRS binding sequence. The findings suggest that employing LAMTOR1 to construct peptides could serve as a promising strategy for bolstering the efficacy of immunotherapy in NSCLC. The discovery and comprehension of how LAMTOR1 inhibits the release of exosomal PD-L1 offer insights into potential therapeutic strategies for improving immunotherapy. It is imperative to conduct further research and clinical trials to investigate the feasibility and efficacy of targeting LAMTOR1 in NSCLC treatment.
Keywords: Exosomes; Immunotherapy; LAMTOR1; Non-small cell lung cancer; PD-L1.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
