

Monocytes Reprogrammed by 4-PBA Potently Contribute to the Resolution of Inflammation and Atherosclerosis
- PMID: 39224974
- PMCID: PMC11424066
- DOI: 10.1161/CIRCRESAHA.124.325023
Monocytes Reprogrammed by 4-PBA Potently Contribute to the Resolution of Inflammation and Atherosclerosis
Abstract
Background: Chronic inflammation initiated by inflammatory monocytes underlies the pathogenesis of atherosclerosis. However, approaches that can effectively resolve chronic low-grade inflammation targeting monocytes are not readily available. The small chemical compound 4-phenylbutyric acid (4-PBA) exhibits broad anti-inflammatory effects in reducing atherosclerosis. Selective delivery of 4-PBA reprogrammed monocytes may hold novel potential in providing targeted and precision therapeutics for the treatment of atherosclerosis.
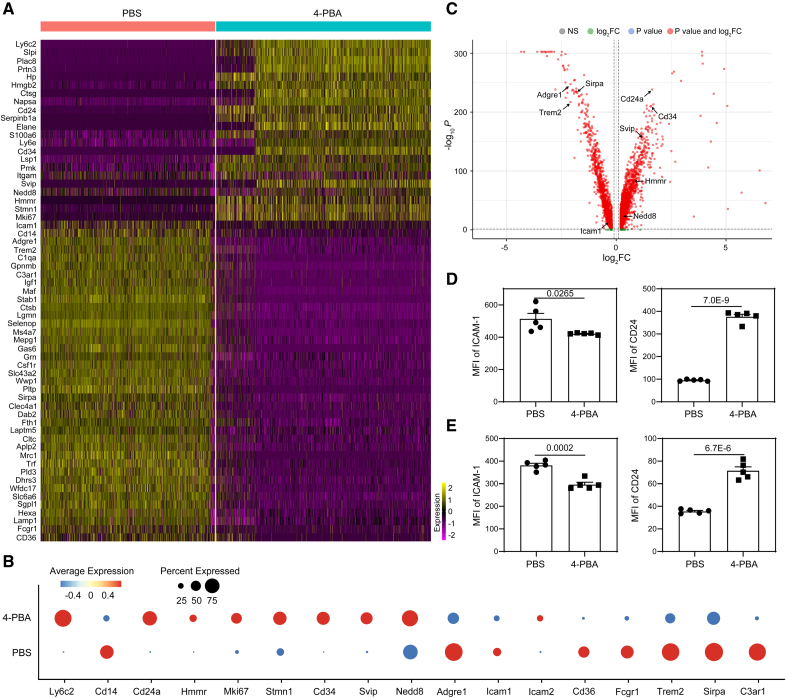
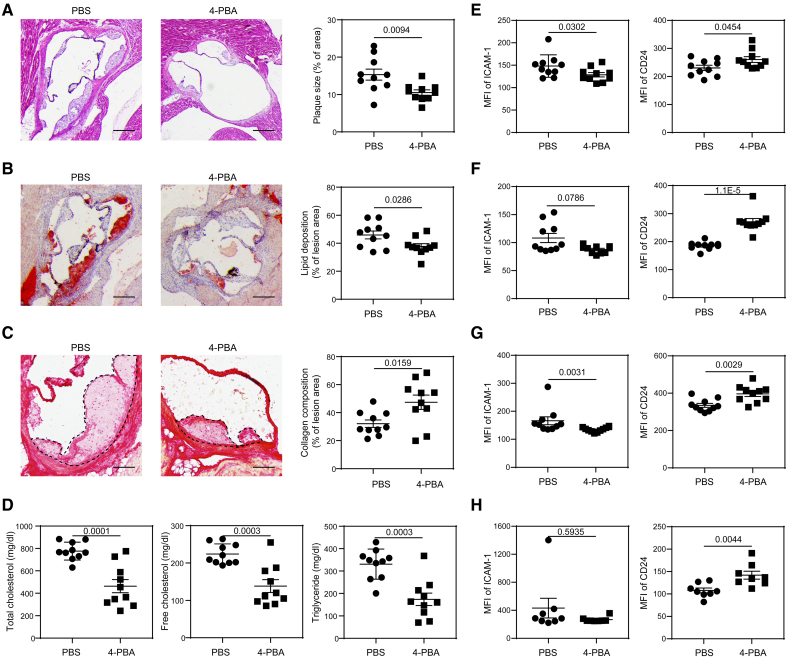
Methods: Systems analyses integrating single-cell RNA sequencing and complementary immunologic approaches characterized key resolving characteristics as well as defining markers of reprogrammed monocytes trained by 4-PBA. Molecular mechanisms responsible for monocyte reprogramming were assessed by integrated biochemical and genetic approaches. The intercellular propagation of homeostasis resolution was evaluated by coculture assays with donor monocytes trained by 4-PBA and recipient naive monocytes. The in vivo effects of monocyte resolution and atherosclerosis prevention by 4-PBA were assessed with the high-fat diet-fed ApoE-/- mouse model with IP 4-PBA administration. Furthermore, the selective efficacy of 4-PBA-trained monocytes was examined by IV transfusion of ex vivo trained monocytes by 4-PBA into recipient high-fat diet-fed ApoE-/- mice.
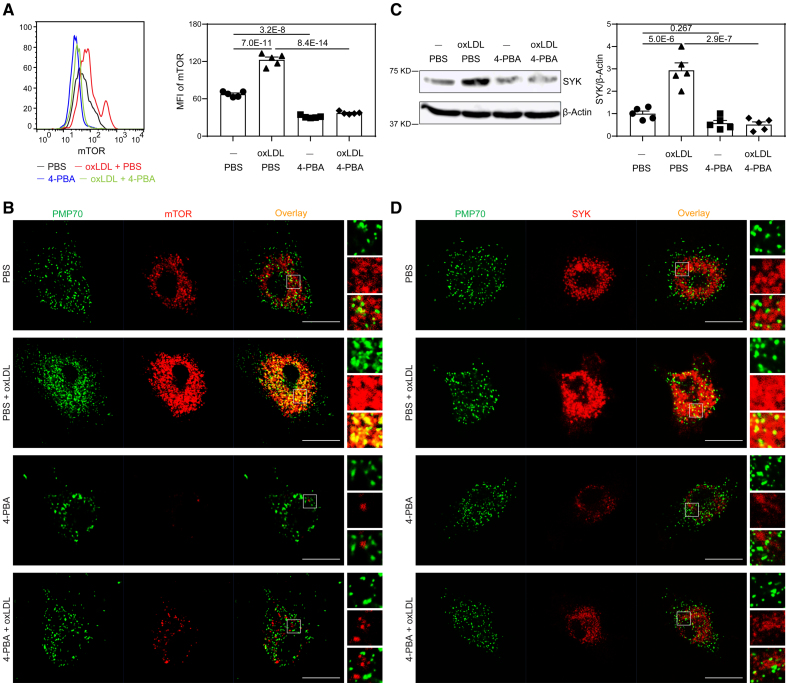
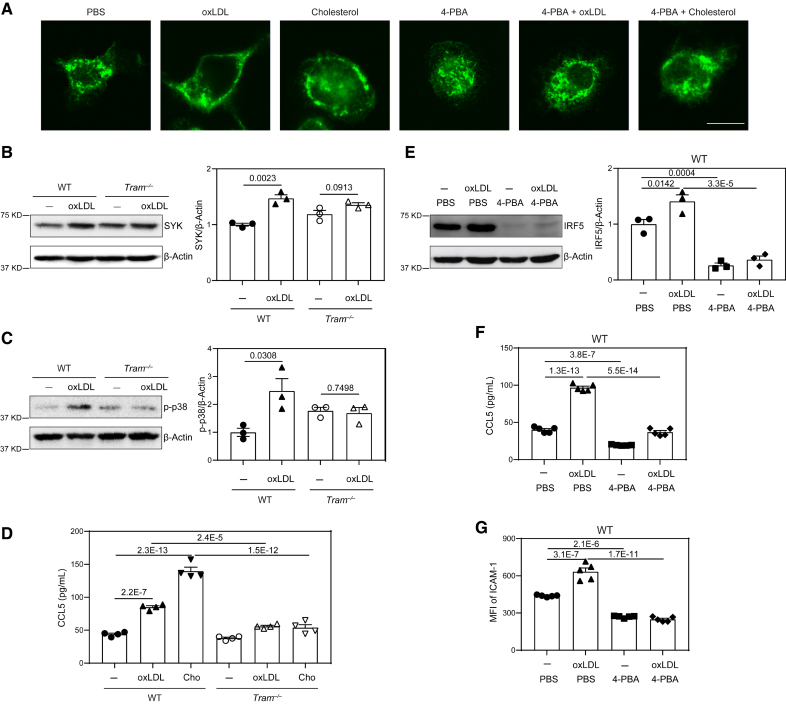
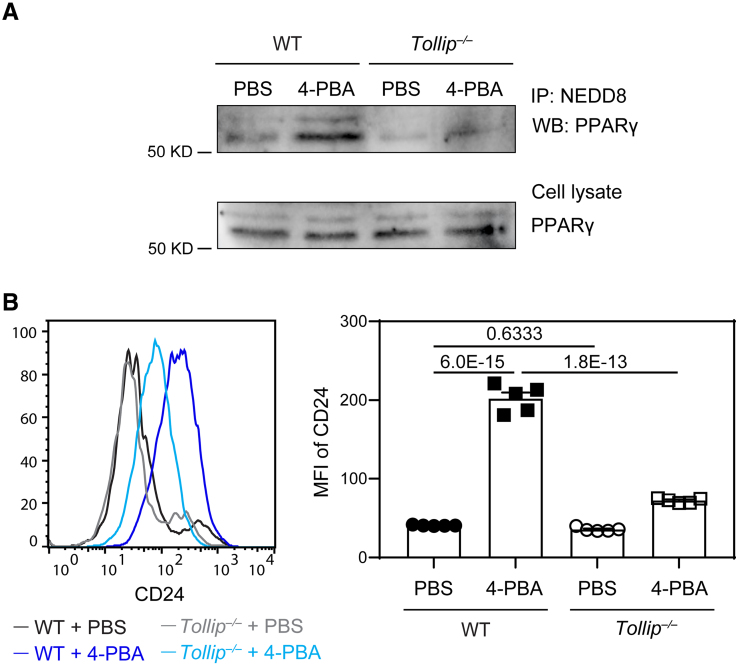
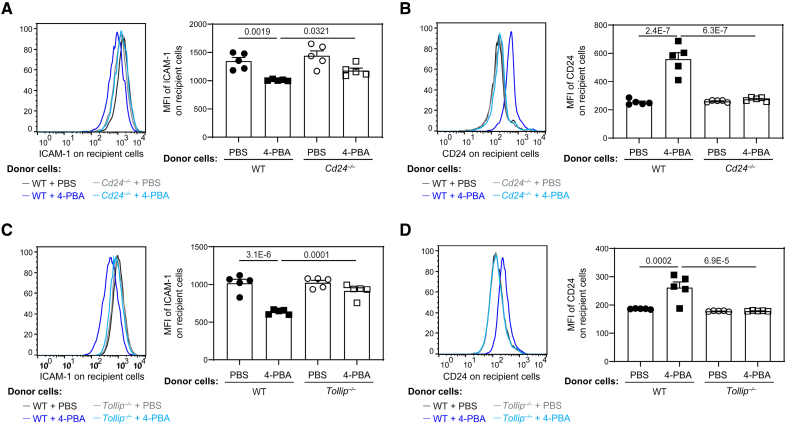
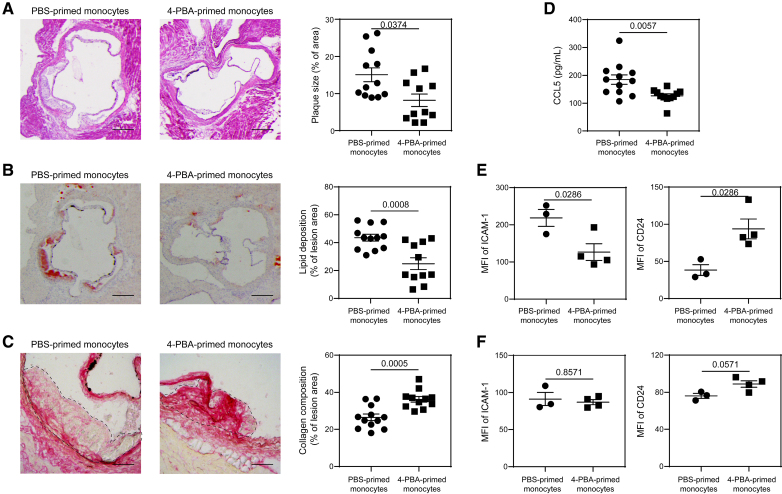
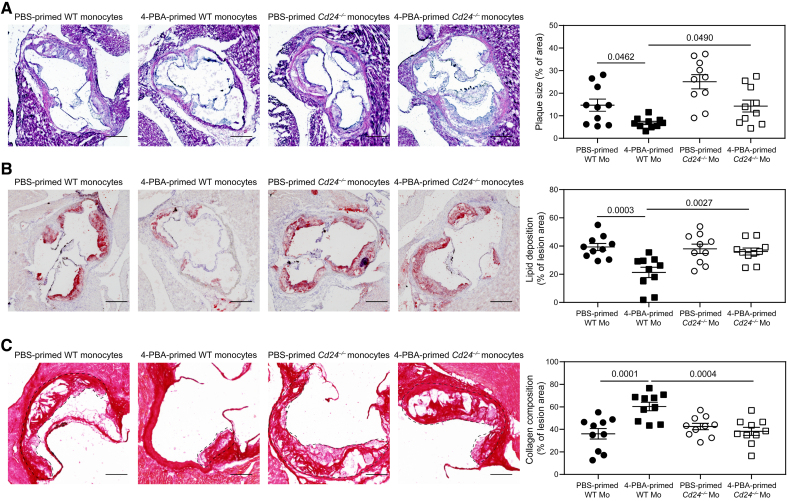
Results: In this study, we found that monocytes can be potently reprogrammed by 4-PBA into an immune-resolving state characterized by reduced adhesion and enhanced expression of anti-inflammatory mediator CD24. Mechanistically, 4-PBA reduced the expression of ICAM-1 (intercellular adhesion molecule 1) via reducing peroxisome stress and attenuating SYK (spleen tyrosine kinase)-mTOR (mammalian target of rapamycin) signaling. Concurrently, 4-PBA enhanced the expression of resolving mediator CD24 through promoting PPARγ (peroxisome proliferator-activated receptor γ) neddylation mediated by TOLLIP (toll-interacting protein). 4-PBA-trained monocytes can effectively propagate anti-inflammation activity to neighboring monocytes through CD24. Our data further demonstrated that 4-PBA-trained monocytes effectively reduce atherosclerosis pathogenesis when administered in vivo.
Conclusions: Our study describes a robust and effective approach to generate resolving monocytes, characterizes novel mechanisms for targeted monocyte reprogramming, and offers a precision therapeutics for atherosclerosis based on delivering reprogrammed resolving monocytes.
Keywords: atherosclerosis; immunity, innate; inflammation; monocytes; therapeutics.
Conflict of interest statement
None.
Figures
Update of
-
Monocytes reprogrammed by 4-PBA potently contribute to the resolution of inflammation and atherosclerosis.bioRxiv [Preprint]. 2023 Oct 23:2023.10.19.563200. doi: 10.1101/2023.10.19.563200. bioRxiv. 2023. Update in: Circ Res. 2024 Sep 27;135(8):856-872. doi: 10.1161/CIRCRESAHA.124.325023. PMID: 37961551 Free PMC article. Updated. Preprint.
References
-
- Ghattas A, Griffiths HR, Devitt A, Lip GY, Shantsila E. Monocytes in coronary artery disease and atherosclerosis: where are we now? J Am Coll Cardiol. 2013;62:1541–1551. doi: 10.1016/j.jacc.2013.07.043 - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous
