

Natural killer cell effector function is critical for host defense against alcohol-associated bacterial pneumonia
- PMID: 39227647
- PMCID: PMC11372167
- DOI: 10.1038/s41522-024-00558-w
Natural killer cell effector function is critical for host defense against alcohol-associated bacterial pneumonia
Abstract
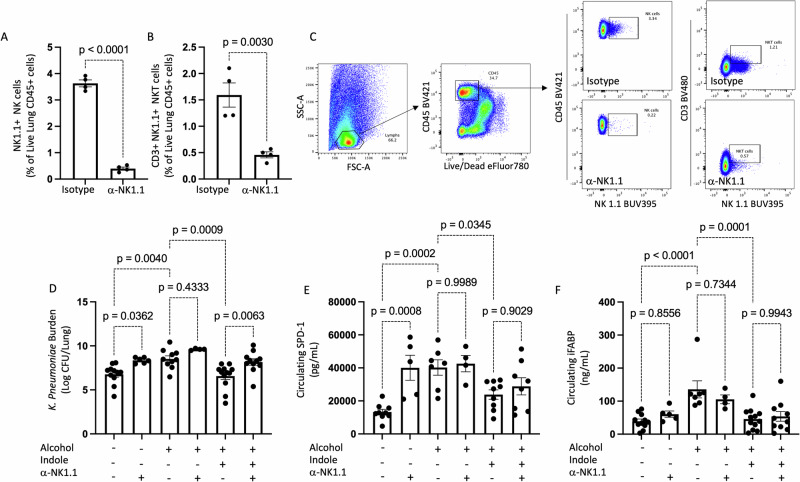
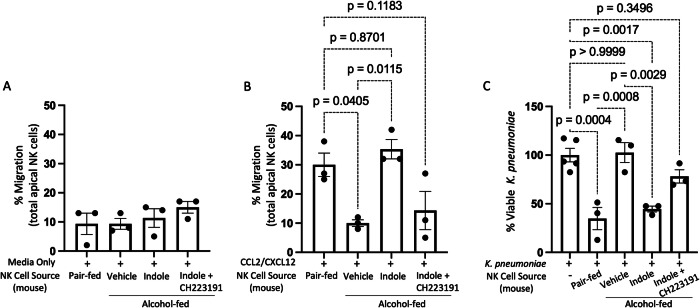
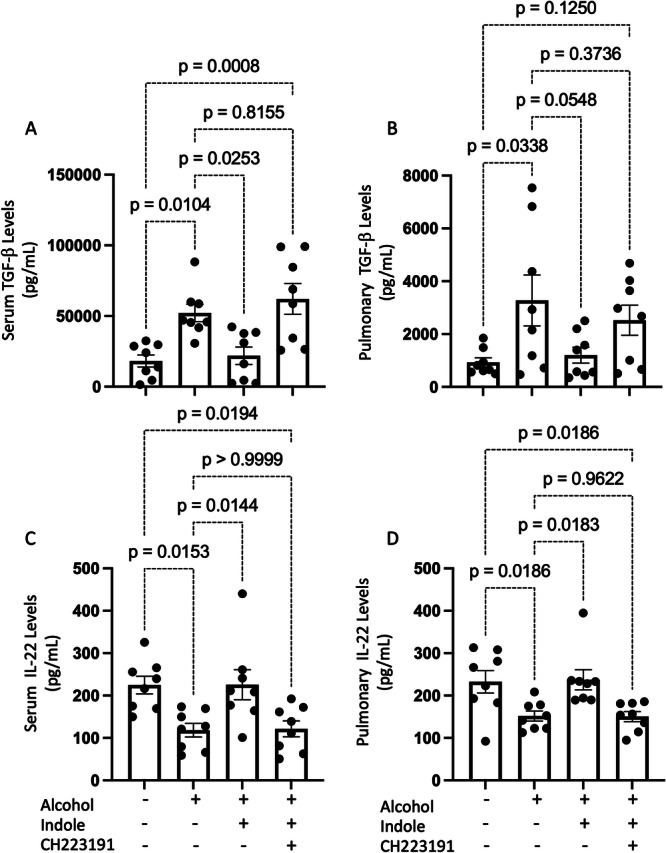
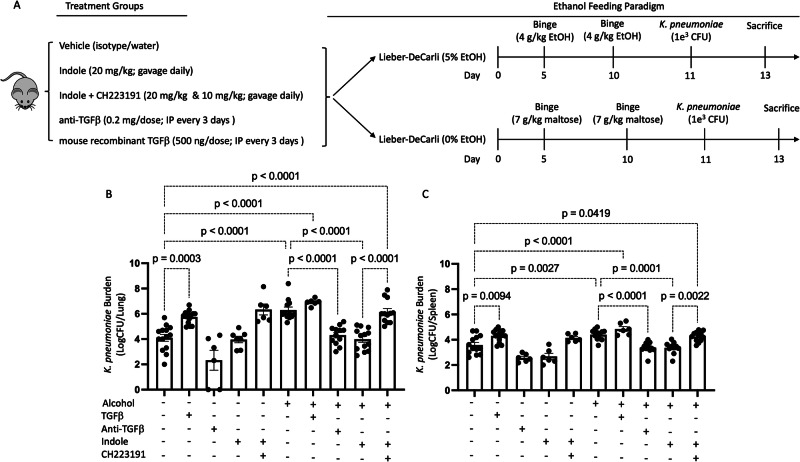
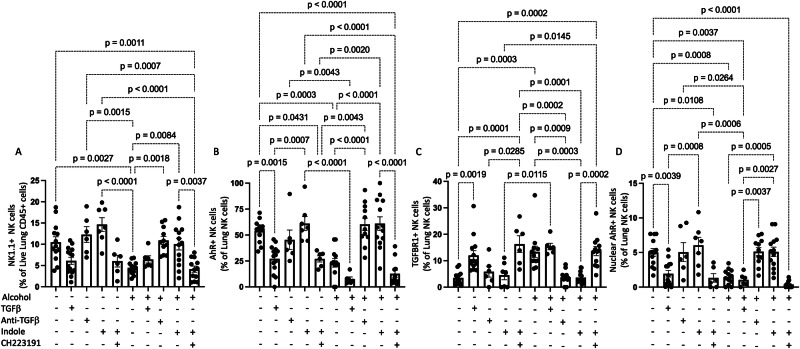
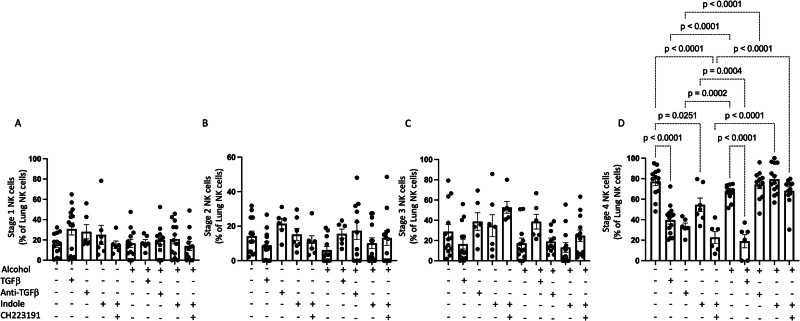
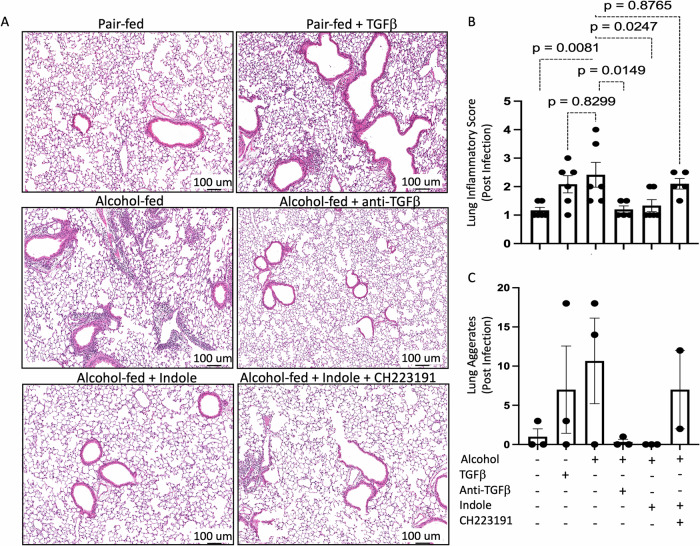
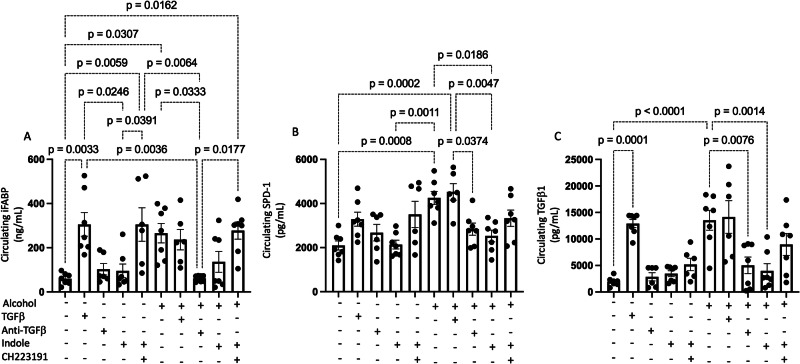
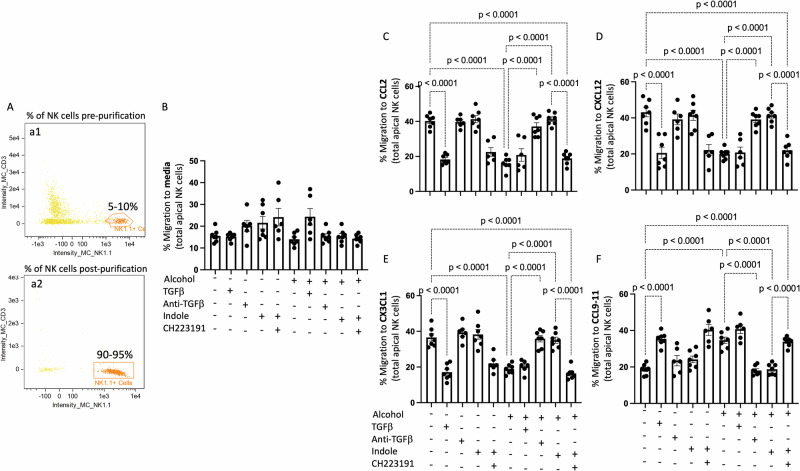
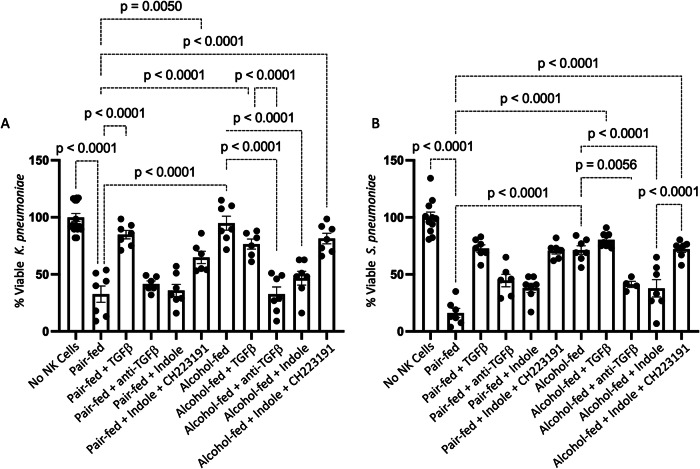
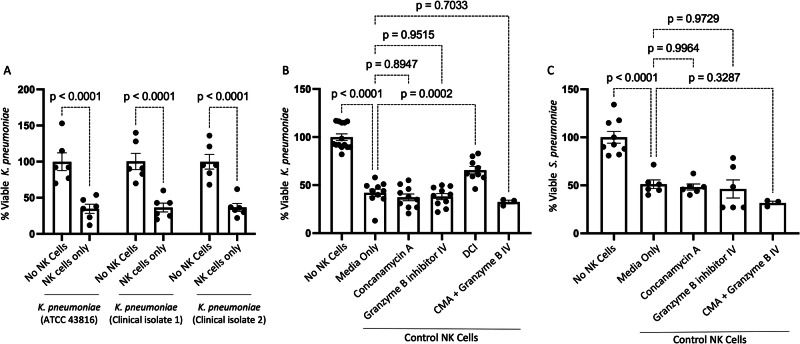
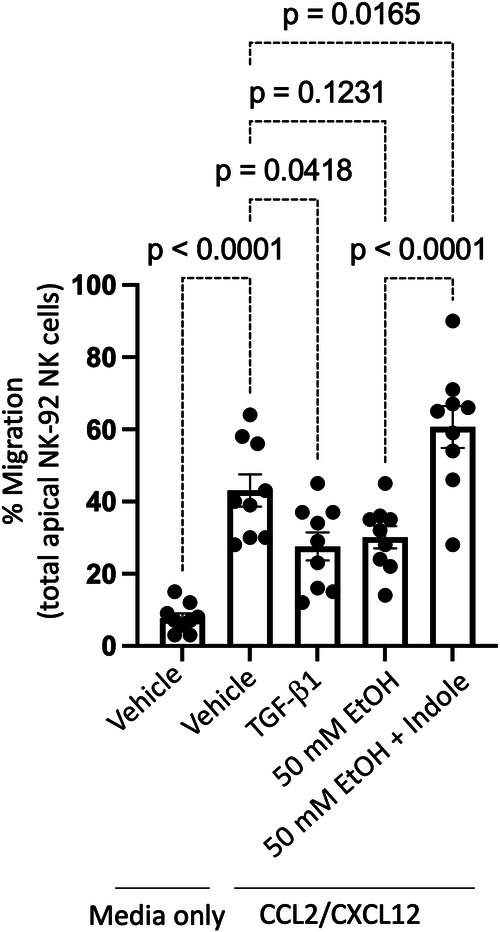
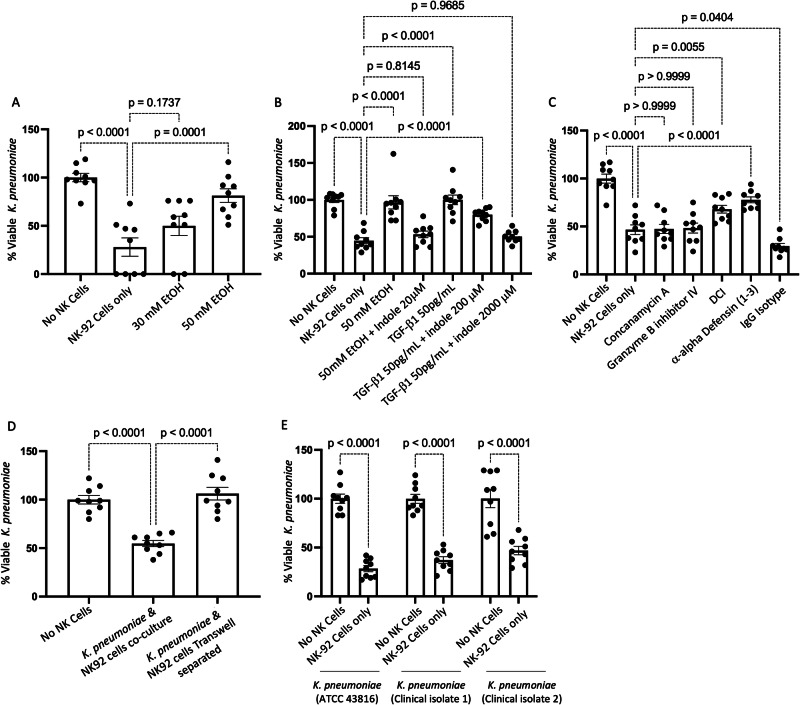
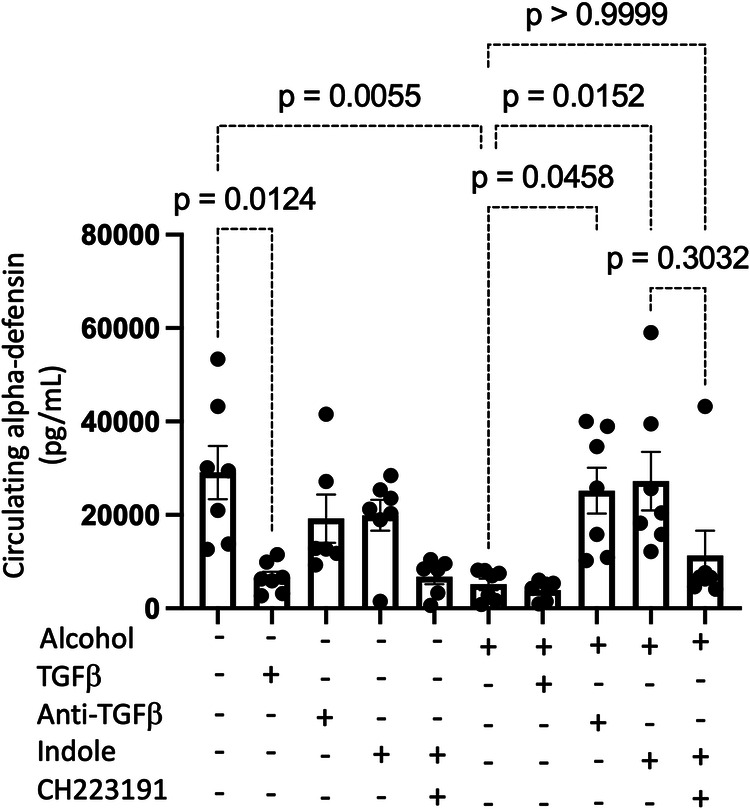
Alcohol use is an independent risk factor for the development of bacterial pneumonia due, in part, to impaired mucus-facilitated clearance, macrophage phagocytosis, and recruitment of neutrophils. Alcohol consumption is also known to reduce peripheral natural killer (NK) cell numbers and compromise NK cell cytolytic activity, especially NK cells with a mature phenotype. However, the role of innate lymphocytes, such as NK cells during host defense against alcohol-associated bacterial pneumonia is essentially unknown. We have previously shown that indole supplementation mitigates increases in pulmonary bacterial burden and improves pulmonary NK cell recruitment in alcohol-fed mice, which were dependent on aryl hydrocarbon receptor (AhR) signaling. Employing a binge-on-chronic alcohol-feeding model we sought to define the role and interaction of indole and NK cells during pulmonary host defense against alcohol-associated pneumonia. We demonstrate that alcohol dysregulates NK cell effector function and pulmonary recruitment via alterations in two key signaling pathways. We found that alcohol increases transforming growth factor beta (TGF-β) signaling while suppressing AhR signaling. We further demonstrated that NK cells isolated from alcohol-fed mice have a reduced ability to kill Klebsiella pneumoniae. NK cell migratory capacity to chemokines was also significantly altered by alcohol, as NK cells isolated from alcohol-fed mice exhibited preferential migration in response to CXCR3 chemokines but exhibited reduced migration in response to CCR2, CXCR4, and CX3CR1 chemokines. Together this data suggests that alcohol disrupts NK cell-specific TGF-β and AhR signaling pathways leading to decreased pulmonary recruitment and cytolytic activity thereby increasing susceptibility to alcohol-associated bacterial pneumonia.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Update of
-
Regulation of Natural Killer Cell TGF-β and AhR Signaling Pathways Via the Intestinal Microbiota is Critical for Host Defense Against Alcohol-Associated Bacterial Pneumonia.Res Sq [Preprint]. 2023 Oct 18:rs.3.rs-3328953. doi: 10.21203/rs.3.rs-3328953/v1. Res Sq. 2023. Update in: NPJ Biofilms Microbiomes. 2024 Sep 3;10(1):79. doi: 10.1038/s41522-024-00558-w. PMID: 37886455 Free PMC article. Updated. Preprint.
References
MeSH terms
Substances
Grants and funding
- R01 HL156952/HL/NHLBI NIH HHS/United States
- R01-DK131990-01/U.S. Department of Health & Human Services | NIH | National Institute of Diabetes and Digestive and Kidney Diseases (National Institute of Diabetes & Digestive & Kidney Diseases)
- R01 DK131990/DK/NIDDK NIH HHS/United States
- P50-AA030407/U.S. Department of Health & Human Services | NIH | National Institute on Alcohol Abuse and Alcoholism (NIAAA)
- IK6-BX005962/U.S. Department of Veterans Affairs (Department of Veterans Affairs)
- IK6 BX005962/BX/BLRD VA/United States
- P50 AA030407/AA/NIAAA NIH HHS/United States
- R24 AA019661/AA/NIAAA NIH HHS/United States
- R25 AA020818/AA/NIAAA NIH HHS/United States
- R01-HL156952/U.S. Department of Health & Human Services | NIH | National Heart, Lung, and Blood Institute (NHLBI)
- F32 AA031180/AA/NIAAA NIH HHS/United States
- R00 AA026336/AA/NIAAA NIH HHS/United States
- R00-AA026336/U.S. Department of Health & Human Services | NIH | National Institute on Alcohol Abuse and Alcoholism (NIAAA)
LinkOut - more resources
Full Text Sources
