

Nucleoporin 153 deficiency in adult neural stem cells defines a pathological protein-network signature and defective neurogenesis in a mouse model of AD
- PMID: 39227892
- PMCID: PMC11373261
- DOI: 10.1186/s13287-024-03805-1
Nucleoporin 153 deficiency in adult neural stem cells defines a pathological protein-network signature and defective neurogenesis in a mouse model of AD
Abstract
Background: Reduction of adult hippocampal neurogenesis is an early critical event in Alzheimer's disease (AD), contributing to progressive memory loss and cognitive decline. Reduced levels of the nucleoporin 153 (Nup153), a key epigenetic regulator of NSC stemness, characterize the neural stem cells isolated from a mouse model of AD (3×Tg) (AD-NSCs) and determine their altered plasticity and gene expression.
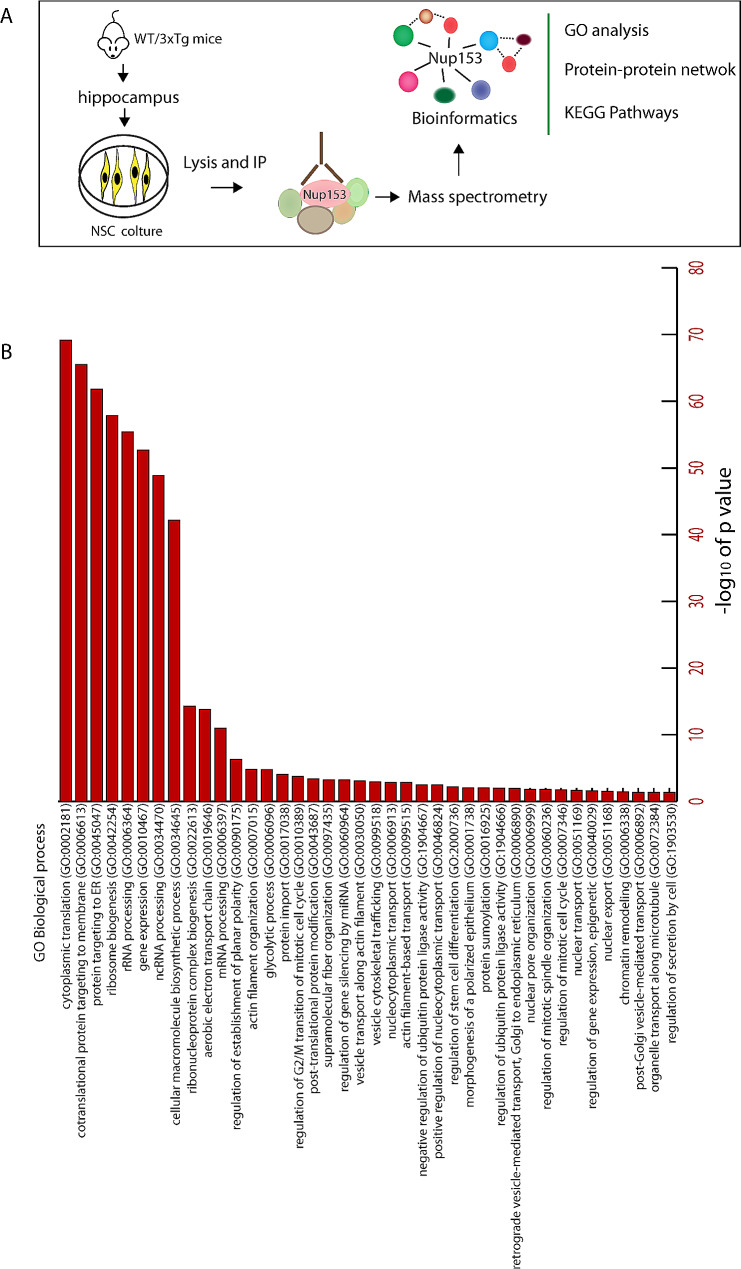
Methods: Nup153-regulated mechanisms contributing to NSC function were investigated: (1) in cultured NSCs isolated from AD and wild type (WT) mice by proteomics; (2) in vivo by lentiviral-mediated delivery of Nup153 or GFP in the hippocampus of AD and control mice analyzing neurogenesis and cognitive function; (3) in human iPSC-derived brain organoids obtained from AD patients and control subjects as a model of neurodevelopment.
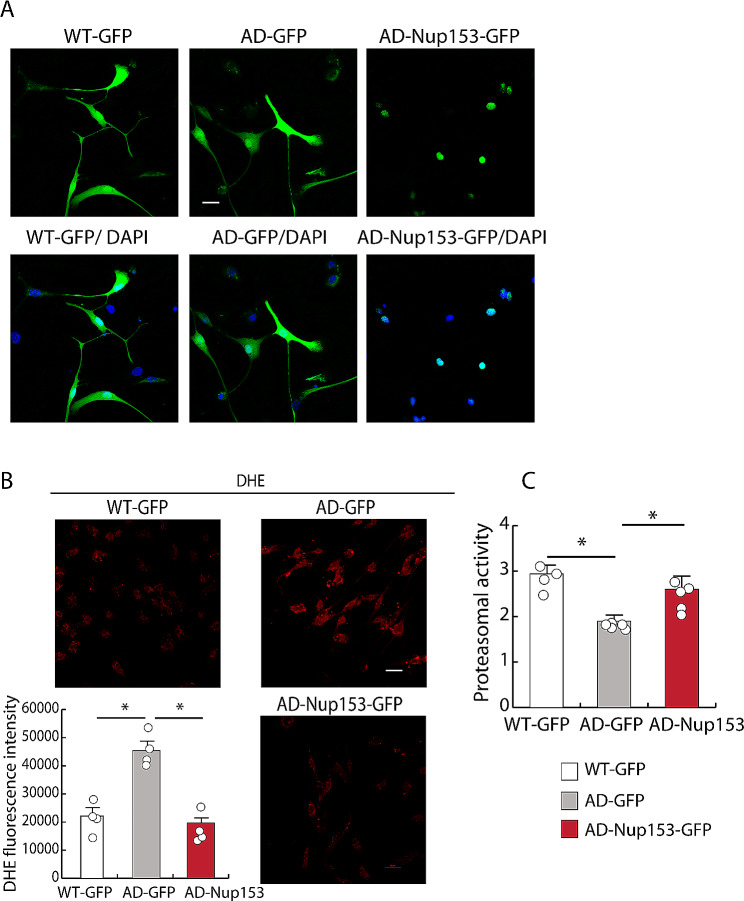
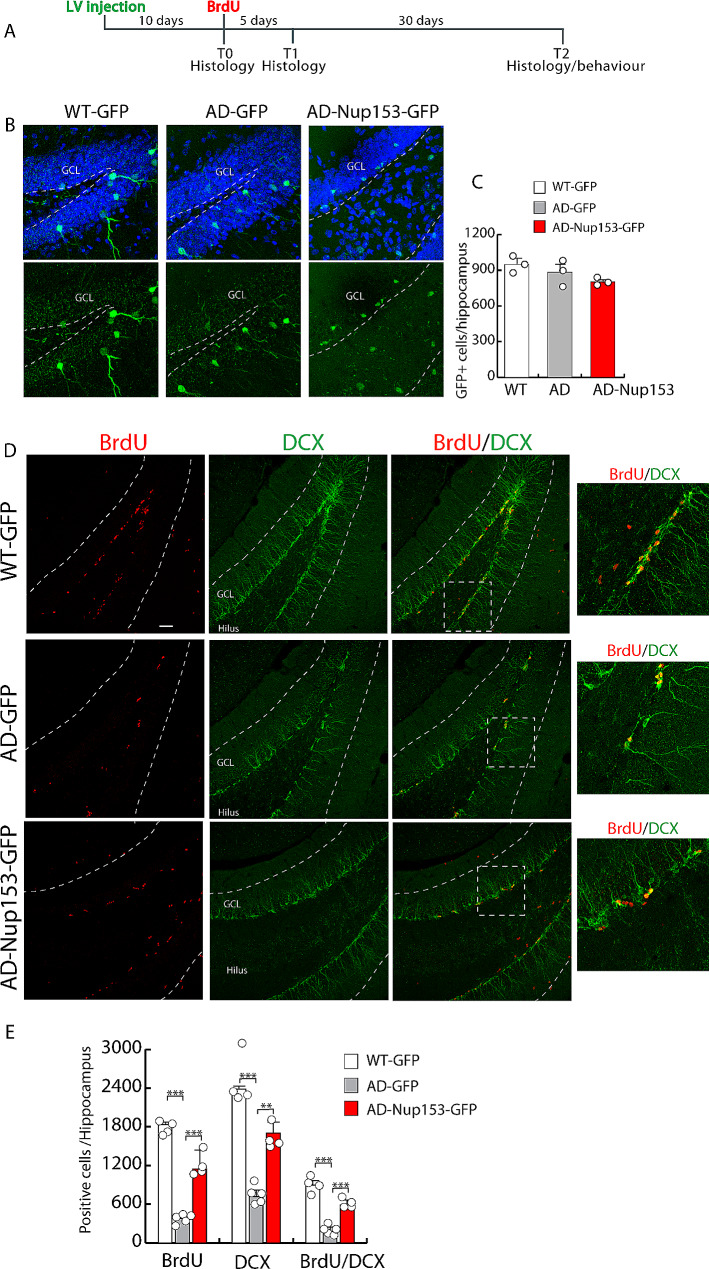
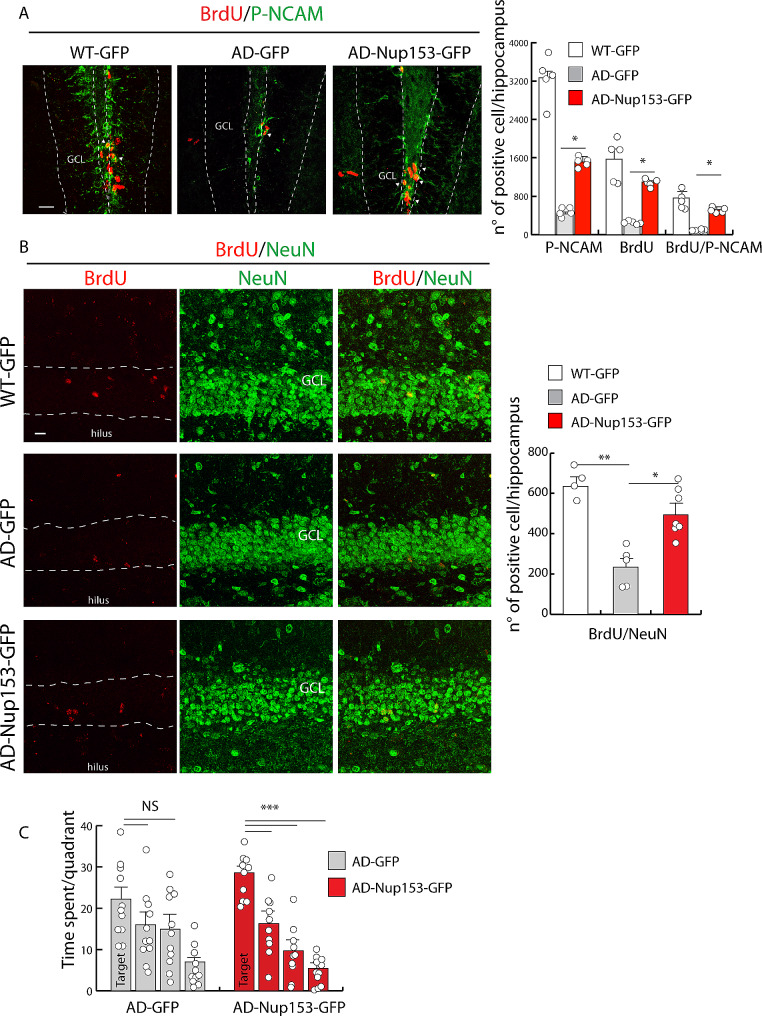
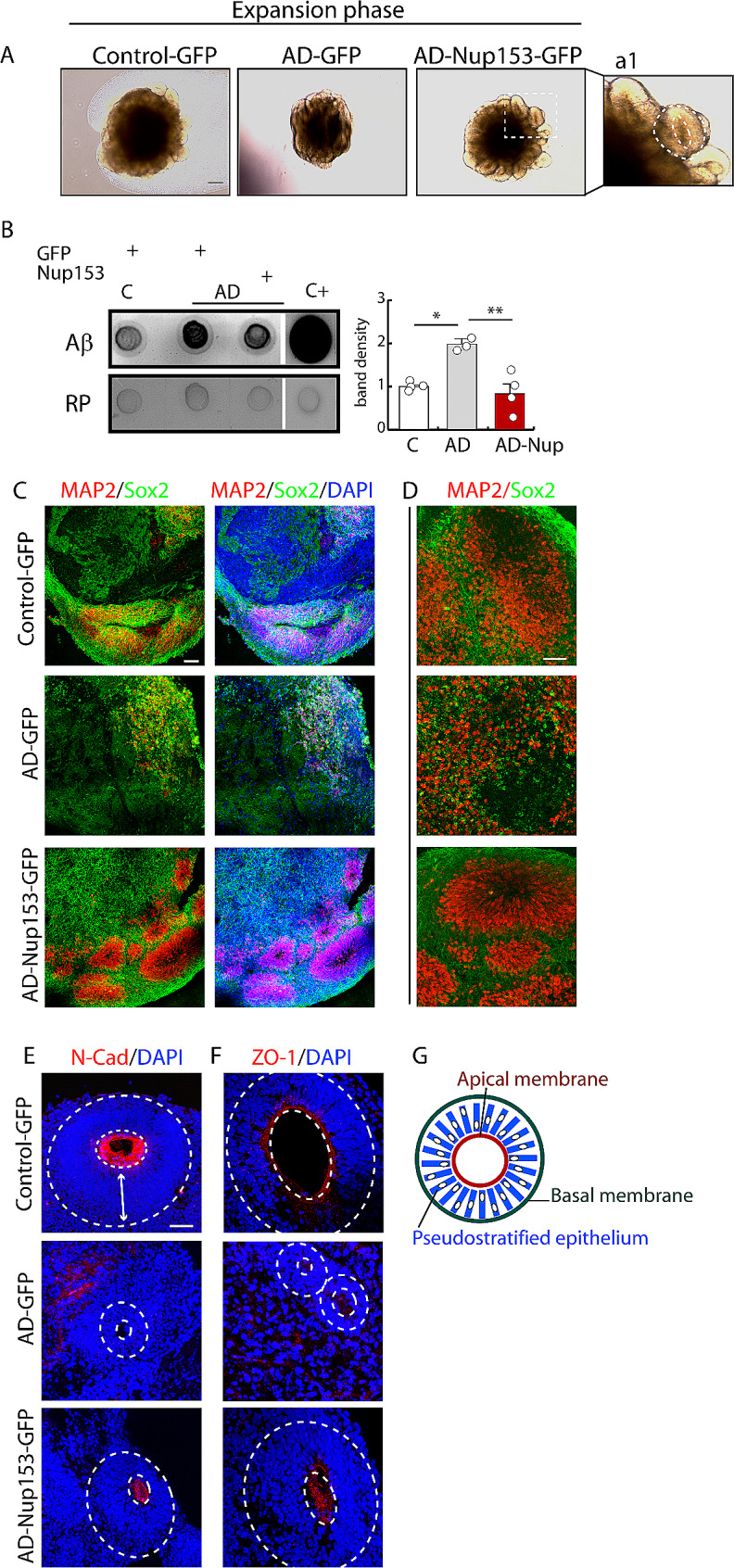
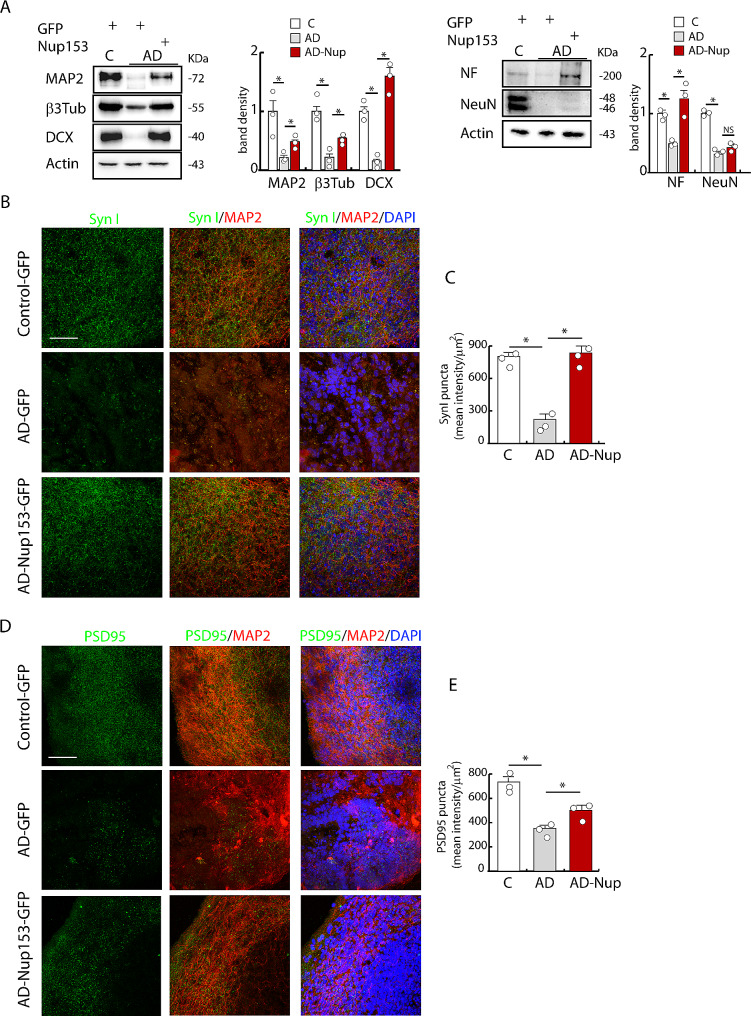
Results: Proteomic approach identified Nup153 interactors in WT- and AD-NSCs potentially implicated in neurogenesis regulation. Gene ontology (GO) analysis showed that Nup153-bound proteins in WT-NSCs were involved in RNA metabolism, nuclear import and epigenetic mechanisms. Nup153-bound proteins in AD-NSCs were involved in pathways of neurodegeneration, mitochondrial dysfunction, proteasomal processing and RNA degradation. Furthermore, recovery of Nup153 levels in AD-NSCs reduced the levels of oxidative stress markers and recovered proteasomal activity. Lentiviral-mediated delivery of Nup153 in the hippocampal niche of AD mice increased the proliferation of early progenitors, marked by BrdU/DCX and BrdU/PSANCAM positivity and, later, the integration of differentiating neurons in the cell granule layer (BrdU/NeuN+ cells) compared with GFP-injected AD mice. Consistently, Nup153-injected AD mice showed an improvement of cognitive performance in comparison to AD-GFP mice at 1 month after virus delivery assessed by Morris Water Maze. To validate the role of Nup153 in neurogenesis we took advantage of brain organoids derived from AD-iPSCs characterized by fewer neuroepithelial progenitor loops and reduced differentiation areas. The upregulation of Nup153 in AD organoids recovered the formation of neural-like tubes and differentiation.
Conclusions: Our data suggest that the positive effect of Nup153 on neurogenesis is based on a complex regulatory network orchestrated by Nup153 and that this protein is a valuable disease target.
Keywords: Alzheimer’s disease; Neurogenesis; Nucleoporin; Organoids.
© 2024. The Author(s).
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Snyder JS, Kee N, Wojtowicz JM. Effects of adult neurogenesis on synaptic plasticity in the rat dentate gyrus. J Neurophysiol. 2001;85(6):2423–31. - PubMed
-
- Kempermann G, Gage FH. Genetic determinants of adult hippocampal neurogenesis correlate with acquisition, but not probe trial performance, in the water maze task. Eur J Neurosci. 2002;16(1):129–36. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
