

Decoding the molecular mechanism of selective autophagy of glycogen mediated by autophagy receptor STBD1
- PMID: 39236246
- PMCID: PMC11406230
- DOI: 10.1073/pnas.2402817121
Decoding the molecular mechanism of selective autophagy of glycogen mediated by autophagy receptor STBD1
Abstract
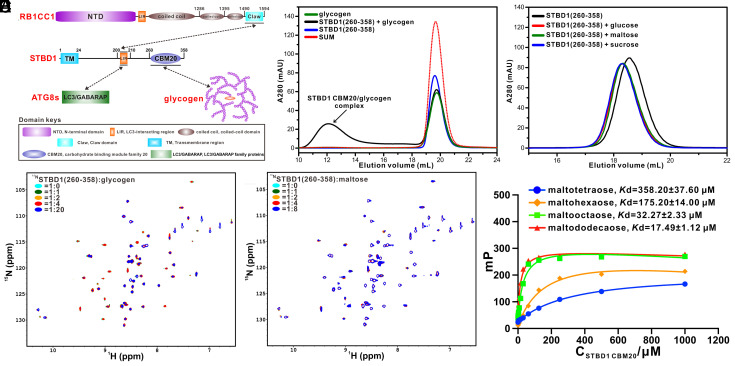
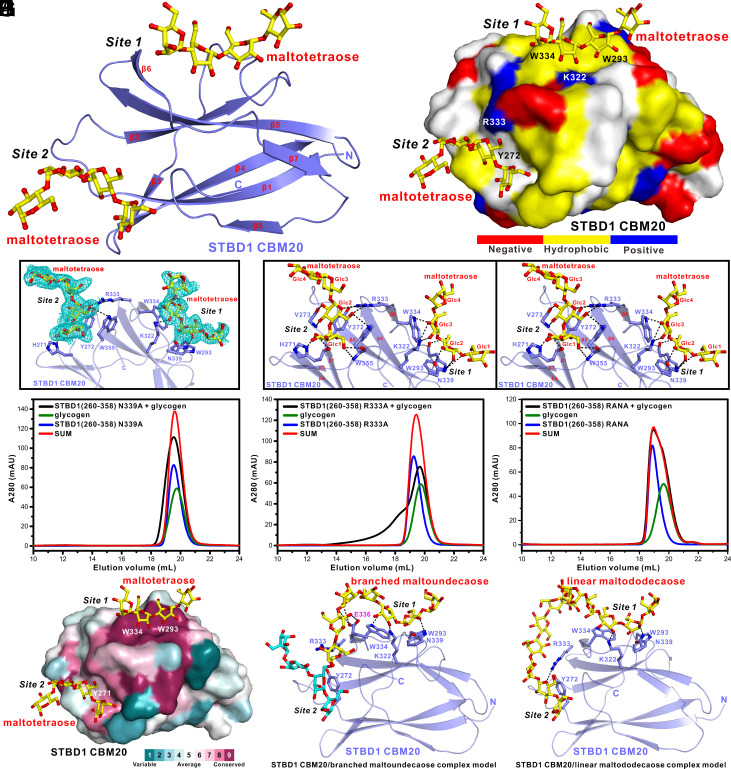
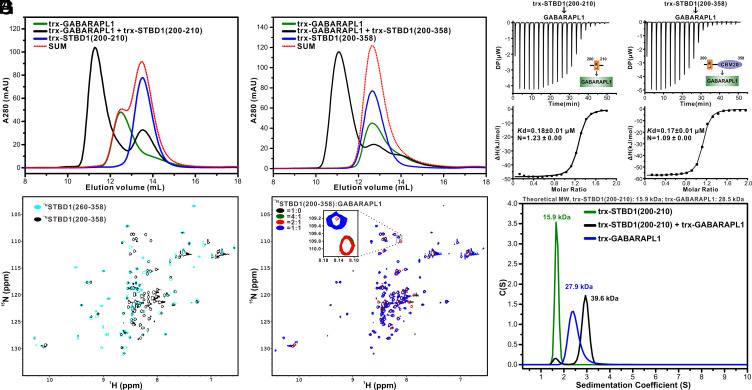
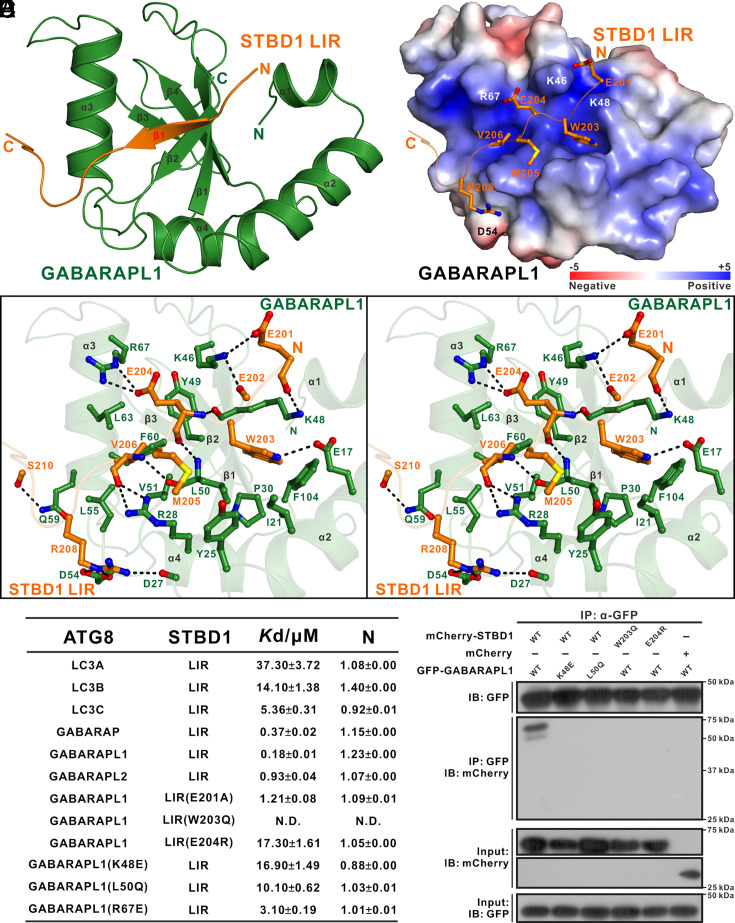
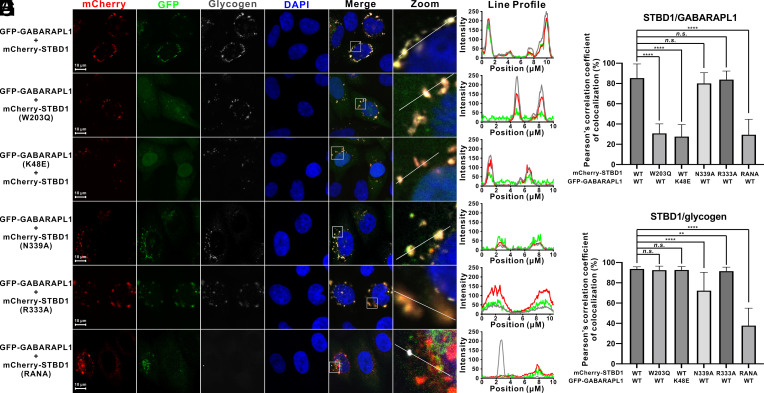
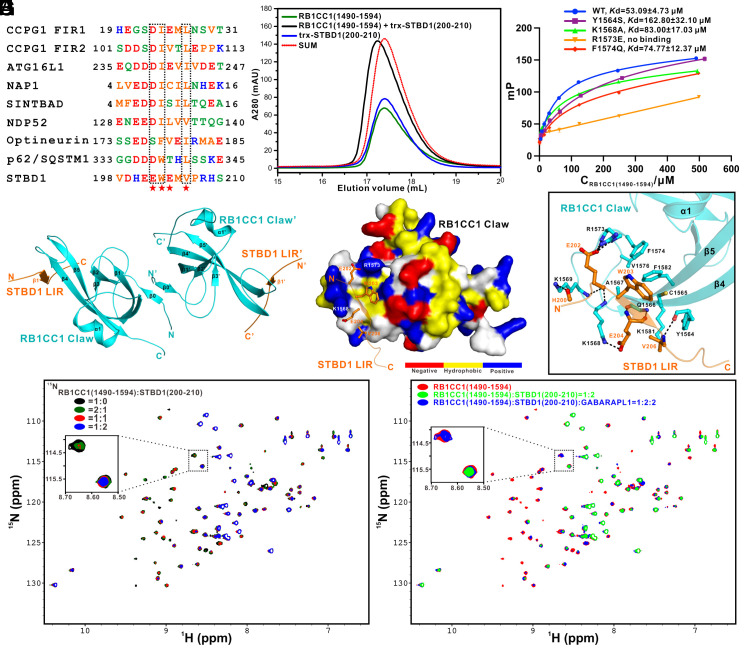
Autophagy of glycogen (glycophagy) is crucial for the maintenance of cellular glucose homeostasis and physiology in mammals. STBD1 can serve as an autophagy receptor to mediate glycophagy by specifically recognizing glycogen and relevant key autophagic factors, but with poorly understood mechanisms. Here, we systematically characterize the interactions of STBD1 with glycogen and related saccharides, and determine the crystal structure of the STBD1 CBM20 domain with maltotetraose, uncovering a unique binding mode involving two different oligosaccharide-binding sites adopted by STBD1 CBM20 for recognizing glycogen. In addition, we demonstrate that the LC3-interacting region (LIR) motif of STBD1 can selectively bind to six mammalian ATG8 family members. We elucidate the detailed molecular mechanism underlying the selective interactions of STBD1 with ATG8 family proteins by solving the STBD1 LIR/GABARAPL1 complex structure. Importantly, our cell-based assays reveal that both the STBD1 LIR/GABARAPL1 interaction and the intact two oligosaccharide binding sites of STBD1 CBM20 are essential for the effective association of STBD1, GABARAPL1, and glycogen in cells. Finally, through mass spectrometry, biochemical, and structural modeling analyses, we unveil that STBD1 can directly bind to the Claw domain of RB1CC1 through its LIR, thereby recruiting the key autophagy initiation factor RB1CC1. In all, our findings provide mechanistic insights into the recognitions of glycogen, ATG8 family proteins, and RB1CC1 by STBD1 and shed light on the potential working mechanism of STBD1-mediated glycophagy.
Keywords: GABARAPL1; RB1CC1; STBD1; glycogen; glycophagy.
Conflict of interest statement
Competing interests statement:The authors declare no competing interest.
Figures
References
-
- Nakatogawa H., Suzuki K., Kamada Y., Ohsumi Y., Dynamics and diversity in autophagy mechanisms: Lessons from yeast. Nat. Rev. Mol. Cell Biol. 10, 458–467 (2009). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
