

FabR, a regulator of membrane lipid homeostasis, is involved in Klebsiella pneumoniae biofilm robustness
- PMID: 39240091
- PMCID: PMC11481535
- DOI: 10.1128/mbio.01317-24
FabR, a regulator of membrane lipid homeostasis, is involved in Klebsiella pneumoniae biofilm robustness
Abstract
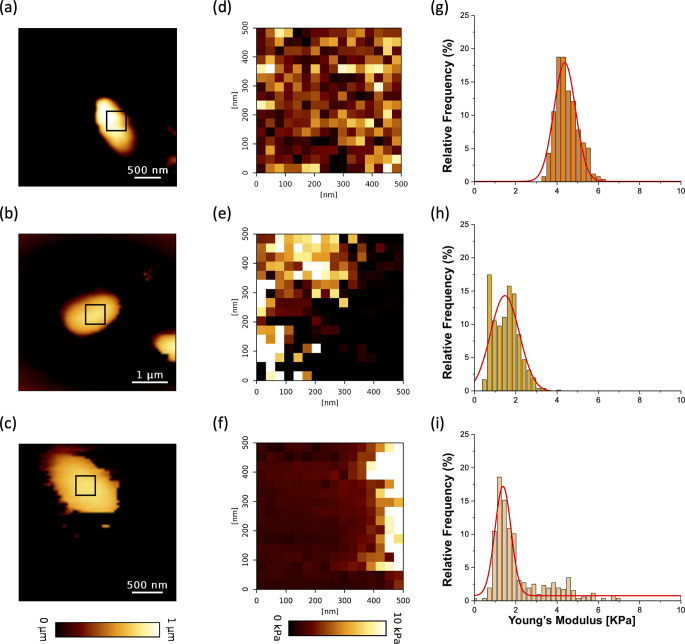
Biofilm is a dynamic structure from which individual bacteria and micro-aggregates are released to subsequently colonize new niches by either detachment or dispersal. Screening of a transposon mutant library identified genes associated with the alteration of Klebsiella pneumoniae biofilm including fabR, which encodes a transcriptional regulator involved in membrane lipid homeostasis. An isogenic ∆fabR mutant formed more biofilm than the wild-type (WT) strain and its trans-complemented strain. The thick and round aggregates observed with ∆fabR were resistant to extensive washes, unlike those of the WT strain. Confocal microscopy and BioFlux microfluidic observations showed that fabR deletion was associated with biofilm robustness and impaired erosion over time. The genes fabB and yqfA associated with fatty acid metabolism were significantly overexpressed in the ∆fabR strain, in both planktonic and biofilm conditions. Two monounsaturated fatty acids, palmitoleic acid (C16:1) and oleic acid (C18:1), were found in higher proportion in biofilm cells than in planktonic forms, whereas heptadecenoic acid (C17:1) and octadecanoic acid, 11-methoxy (C18:0-OCH3) were found in higher proportion in the planktonic lifestyle. The fabR mutation induced variations in the fatty acid composition, with no clear differences in the amounts of saturated fatty acids (SFA) and unsaturated fatty acids for the planktonic lifestyle but lower SFA in the biofilm form. Atomic force microscopy showed that deletion of fabR is associated with decreased K. pneumoniae cell rigidity in the biofilm lifestyle, as well as a softer, more elastic biofilm with increased cell cohesion compared to the wild-type strain.IMPORTANCEKlebsiella pneumoniae is an opportunistic pathogen responsible for a wide range of nosocomial infections. The success of this pathogen is due to its high resistance to antibiotics and its ability to form biofilms. The molecular mechanisms involved in biofilm formation have been largely described but the dispersal process that releases individual and aggregate cells from mature biofilm is less well documented while it is associated with the colonization of new environments and thus new threats. Using a multidisciplinary approach, we show that modifications of bacterial membrane fatty acid composition lead to variations in the biofilm robustness, and subsequent bacterial detachment and biofilm erosion over time. These results enhance our understanding of the genetic requirements for biofilm formation in K. pneumoniae that affect the time course of biofilm development and the embrittlement step preceding its dispersal that will make it possible to control K. pneumoniae infections.
Keywords: Klebsiella pneumoniae; atomic force microscopy; biofilm robustness; membrane fatty acid.
Conflict of interest statement
The authors declare no conflict of interest.
Figures








References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
